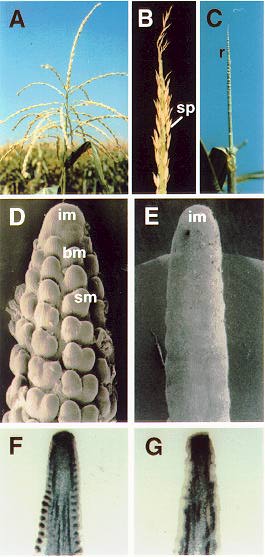
barren inflorescence2 (bif2) mutants are characterized by a reduction in the number of spikelets in the tassel and ear and a reduction in the number of lateral branches on the tassel (Fig. 1B). An EMS allele, bif2-2354, and a Mu-induced allele, bif2-47330, have previously been described (Briggs, S and Johal, G, MNL 66:51 1992; Neuffer, MG and Briggs, S, MNL 68:28, 1994). We describe two new alleles, isolated from lines containing active Mutator (Mu) elements, which were kindly given to us by Paul Chomet and Guri Johal. The mutants fail to complement bif2 and hence are designated, bif2-1606 and bif2-70. The new alleles have a more severe phenotype than bif2-2354. Usually, a thin bare unbranched rachis (inflorescence stem) forms in place of the tassel (Fig. 1C). Spikelets that form infrequently are usually single instead of paired and have elongated pedicels. Occasionally, one or two lateral branches, with a limited number of spikelets, are produced. The ear is similarly affected : ear shoots form but when the husk leaves are opened, only a bare rachis is present (not shown). A few florets may be found at the base of the ear or in small patches on the rachis. In addition, the ear is almost always fasciated. In contrast to the dramatic effect on inflorescence development, vegetative development appears normal. We propose that bif2 is required for the formation of branches and spikelets. Infrequent branch and spikelet formation may be due to redundancy with other barren inflorescence loci.
Scanning electron microscopy was used to determine why bif2-1606 mutants do not produce branches and spikelets. The first step in normal inflorescence development is the formation of branch meristems by the inflorescence meristem (Fig. 1D). Branch meristems at the base of the tassel reiterate the main spike to produce long branches. Branch meristems on the main spike and the lateral branches produce two spikelet meristems, each of which produces two floret meristems (Fig. 1D). In bif2-1606 inflorescences, no branch meristems were produced by the inflorescence meristem (Fig. 1E). Ripples or undulations on the surface of the rachis may represent the suppressed bract primordia which normally subtend branch meristems. Therefore, we propose that bif2 mutants do not produce branches and spikelets because they are blocked in inflorescence development before the production of branch meristems.
This conclusion was confirmed by RNA in situ hybridization using kn1 as a marker for meristematic tissue. In normal inflorescences, kn1 is expressed in the inflorescence, branch, spikelet and floret meristems (Jackson, D et al., Development 120:405-413, 1994; Fig. 1F). In bif2-1606 inflorescences, kn1 was expressed in the inflorescence meristem but was not expressed on the flanks of the inflorescence meristem where branch meristems initiate (Fig. 1G). Similar results were obtained with bif2-70. This confirms that bif2 mutants do not produce branch meristems. Whether bif2 is required for the initiation or the maintenance of branch meristems is currently being investigated.
Our results show that bif2 plays a fundamental role in formation of branch meristems which is the first step in the production of florets. To determine the molecular mechanism by which it acts we are currently attempting to clone bif2 by transposon tagging.
Figure 1. A) Normal tassel. B) bif2-2354 tassel with no branches and a reduced number of spikelets on the main spike. C) bif2-1606 tassel with no branches or spikelets. D) SEM of a normal tassel. E) SEM of a bif2-1606 tassel showing that branch meristems are not produced. F) RNA in-situ hybridization showing kn1 expression (blue) in a normal ear. G) RNA in-situ hybridization of kn1 expression in a bif2-1606 ear.
Abbreviations : sp, spikelet, r, rachis, im, inflorescence meristem, bm, branch meristem, sm, spikelet meristem.
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}