Discovery and primary study on photoperiod-sensitive male sterility
in maize
--Zhou, H; Deng, Y; Tian, Z
The photoperiod-sensitive male sterility in maize first reported here was discovered by us in 1995. Over 3 years and 3 sites, we found that expression of fertility in maize line CA507 was not stable and had some correlation with the photoperiod. When planted in short-photoperiod conditions, it expressed male sterility. Otherwise it was male fertile under long-photoperiod conditions. The threshold of light length may be 14-15 hours. Preliminary study indicated that over 2 recessive genes underlaid this trait.
Since the first photoperiod-sensitive male sterility in rice was discovered by Shi (Hubei Agric. Sci. 7:1-3, 1981), many photoperiod-sensitive types of male sterility in other crops have been found one after another, such as in barley (Wang and Cheng, Hubei Agric. Sci. 11:14-16, 1991), in wheat (He et al., Hunan Agric. Sci. 5:1-3, 1992). Photoperiod-sensitive male sterility has a unique role in hybrid seed production, especially for self-pollinated crops. More and more facts support the view that photoperiod and temperature co-act in male sterility. Generally, if photoperiod acts dominantly we call it photoperiod-sensitive male sterility and vice versa.
He et al. (Crops 2:1-2, 1995) first reported their finding of temperature-sensitive male sterility in maize in 1995. We have been looking for photoperiod-sensitive maize for several years, because we believe that photoperiod sensitive material should exist in maize as in other crops. Photoperiod-sensitive male sterility should be more stable than temperature-sensitive because photoperiod compared with temperature is more steady and predictable among different years, and therefore has a great advantage in practical application.
In the winter of 1994, we found in Nanbin farm, Sanya city, Hainan province that all the 38 plants of inbred line CA507 displayed male sterility. The anthers of the inbreds didn't shed and had no normal pollen in them but a few irregular grains which could not be dyed to dark-blue color when treated with KI. All the plants displayed complete male sterility. From then on, we have conducted a series of studies on this character.
CA507, and F1s of CA507 crossed with other five normal inbreds, were planted in Beijing and in Sanya respectively in 1994 and 1995. CA507 was grown both in long and in short photoperiod conditions in Wuhan University in 1996. Three F2s and one BC1 were planted in Sanya and Beijing in 1996 for the evaluation of gene acting mode. The fertility of pollen was determined through KI dyeing under microscope.
Seeds from the same selfed ear of CA507 were grown both in Sanya (latitude 18.3 N) and in Beijing (latitude 40 N) (see details in Table 1). While all the 38 plants displayed complete male sterility in Sanya, only 10 plants were male sterile and the other 10 plants were male fertile in Beijing. In order to detect whether this fertility difference resulted from the difference of photoperiod or temperature between the two locations, we arranged another experiment in Wuhan University under precisely controlled environment (Table 2) and collected the related temperature data of the three locations during the growing season (Fig. 1, 2, 3).
Table 1. Fertility statistics of CA507 under different photoperiods
in Sanya and Beijing (1994-1995).
| Location | Total plants | Fertile | Sterile | Light hours/day |
| 94Sanya | 38 | 0 | 38 | 11-12 |
| 95Beijing | 20 | 10 | 10 | 14.5-15.5 |
Two sets of CA507 seeds were grown respectively in controlled culture rooms with 11 and 17.5 light hours per day in Wuhan University in 1996. Among the 19 plants treated with 11 light hours per day, 18 plants displayed male sterility and 1 plant displayed partial male fertility in a few tassel branches. However, under the 17.5 light hours condition, all the 18 tested plants were male fertile.
Table 2. Reaction of CA507 to different photoperiod in controlled conditions
in Wuhan (1996).
| Light-hours | Total plants | Fertile | Partial fertile | Sterile |
| 11 | 19 | 0 | 1 | 18 |
| 17.5 | 18 | 18 | 0 | 0 |
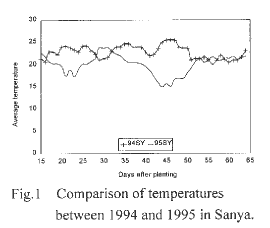
Figure 1. Comparison of temperatures between 1994 and 1995 in Sanya.
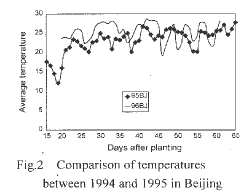
Figure 2. Comparison of temperatures between 1994 and 1995 in Beijing.
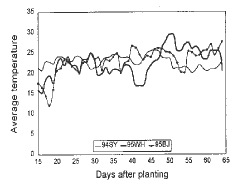
Figure 3. Comparison of temperatures among Sanya, Wuhan and Beijing.
From Figure 1, though the temperature difference between 1994 and 1995 was remarkable in Sanya during 20-25 days and 40-50 days after planting, the fertility of the test plants didn't change much. Temperature didn't vary much between 1995 and 1996 in Beijing (Fig. 2) and between Beijing and Wuhan in 1995 (Fig. 3). From the results of our experiments in the 3 sites, we could conclude that the main factor affecting fertility was photoperiod, not temperature.
In summary, we have the following primary results: 1) CA507 displayed male sterility in short photoperiod conditions and male fertility in long photoperiod conditions, which indicated that CA507's fertility was photoperiod-sensitive; 2) the 14.5-15.5 light hours in Beijing was perhaps the threshold for fertility transition of CA507, which accounted for the coexistence of sterile and fertile type of CA507 when grown in Beijing. If we extended the light time to 17.5 hours per day, all the test plants displayed male fertility.
In Table 3, all the 304 F1 plants in the first 4 crosses showed male fertility both in Beijing and in Sanya, but 3 partial fertile plants in the 17 plants in cross CA507 x Duo4-3 in Sanya. This indicated that CA507's photoperiod-sensitivity may be controlled by recessive genes. But we used just 5 inbreds to test it; this needs further verification with precise genetic design.
Table 3. Phenotypes in F1 (1995-1996).
| Crosses | Sanya | Beijing | |||
| TP | FP | PFP | TP | FP | |
| CA507 x Huangzhao4 | 37 | 37 | 0 | 51 | 51 |
| CA507 x 1007 | 38 | 38 | 0 | 70 | 70 |
| CA507 x Zhi26-5 | 37 | 37 | 0 | 68 | 68 |
| CA507 x 1006 | 29 | 29 | 0 | 46 | 46 |
| CA507 x Duo4-3 | 17 | 14 | 3 | 69 | 69 |
In F2 and BC1 (see detail in Table 4), we found that all the F2 plants of CA507 x Huangzhao4 and CA507 x 1007 were male fertile both in Beijing and in Sanya. However, there were 1 male sterile plant in 20 plants of F2 (CA507 x 1006) and 1 in 17 plants of BC1 (CA507 x (CA507 x 1007)). We reckoned that over 2 pairs of genes controlled CA507's sensitivity to photoperiod. But the population was not big enough to identify the genetic mode precisely. Further study should be done on this.
Table 4. Phenotypes in F2 and backcross (1996).
| Crosses | Generation | Sanya | Beijing | |||
| TP | FP | SP | TP | FP | ||
| (CA507 x Huangzhao4) | F2 | 27 | 27 | 0 | 38 | 38 |
| (CA507 x 1007) | F2 | 29 | 29 | 0 | 27 | 27 |
| (CA507 x 1006) | F2 | 20 | 19 | 1 | ||
| CA507 x (CA507 x 1007) | BC1 | 17 | 16 | 1 |
Through several years study, we obtained the following primary results:
1) CA507's fertility was photoperiod-sensitive; 2) 14.5-15.5 light hours
was perhaps the fertility/sterility transition period for CA507; 3) over
2 pairs of recessive genes control CA507's character of photoperiod sensitivity;
4) the genetic mechanism of this phenomenon still needs further study.
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}