Mutator-induced anomalous alleles of the blue fluorescent1
(bf1)
locus in maize
--Robertson, DS
The original blue fluorescent1 (Bf1-R) mutant allele was described by Teas and Anderson (Proc. Nat. Acad. Sci. USA 37:645, 1951). This mutant accumulates the ?-glucoside of anthranilic acid, anthranilic acid and an unknown compound due to altered inhibition of anthranilic acid metabolism (Singh & Widholm, Biochem. Genet. 13:357, 1975). Homozygous mutant seedlings, and the anthers of plants homozygous or heterozygous for the mutant allele, fluoresce blue under ultraviolet light. Mutant alleles have been designated Bf1, and normal alleles bf1 (see Table 1 for the description of symbols used in this note to designate the alleles and phenotypes involving this locus.)
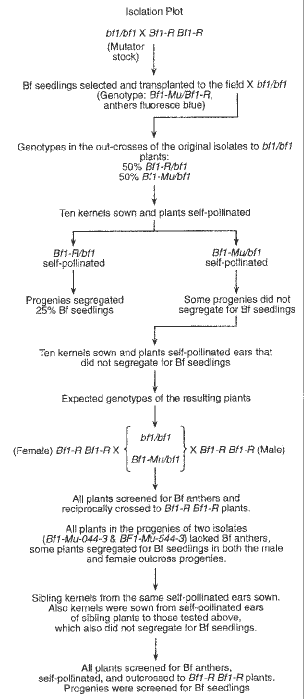
From seeds produced in an isolation plot (see schematic in Fig. 1) in which the female parent plants were active Mutator plants and the male parents were homozygous for the Bf-R allele, 214 seedlings with the Bf phenotype were recovered (genotype = Bf1-Mu/Bf1-R). These seedlings were transferred to the field. At maturity their anthers were checked for pollen abortion and the Bf phenotype and they were then pollinated by bf1 bf1 plants. Half of the kernels from these ears were expected to be heterozygous for the Bf1-R allele (genotype Bf1-R/bf1) and half heterozygous for the Bf1-Mu allele (an isolate, genotype = Bf1-Mu/bf1). For about 50% of Mutator-induced mutants generally, mutability can be used to distinguish the induced mutant from the standard mutant allele. However, mutability could not be utilized for this purpose with Bf1-Mu mutants because the intensity of the blue fluorescence made it impossible to observe the small revertant sectors, which are characteristic of Mutator-induced mutants. Thus, another criterion had to be employed to distinguish the plants with the Mutator-induced mutant alleles (or aberrations) from those with the standard mutant allele. The criterion used was to screen progenies of self-pollinated plants from the outcrosses of the Bf1-Mu/Bf1-R plants as females to homozygous bf1 stocks (resulting genotypes = Bf1-Mu/bf1 & Bf1-R/bf1) for absence or reduced numbers of Bf seedlings.
The following are some of the events that might be responsible for such a phenomenon and possible observations that would be expected if the events occurred: 1) Induced deletions involving the bf1 locus, which were not male transmissible. Plants heterozygous for such deletions when outcrossed as males to Bf1-R tester plants would have no Bf seedlings in the outcross progenies but 50% or less Bf seedlings would be expected when the plants were crossed as females to the same tester stocks [Putative Mutator-induced deletions involving the yg2 locus that were not male transmissible were found by Robertson and Stinard (Genetics 115:353, 1987)]. Also pollen abortion might be expected in plants heterozygous for such deletions [See Robertson (Genome 37:433, 1996) for the description of a Bf1-Mu deletion characterized by the segregation of abortive pollen grains]. 2) The event, when homozygous, could result in the failure of the endosperm to develop. If this occurred, self-pollinated ears of heterozygous plants would not have normal seed set. 3) The homozygous event could interfere with the development of the germ. Such an event would result in the segregation of germless kernels on the ears of self-pollinated heterozygous plants. 4) The event could affect the ability of homozygous kernels to germinate. If germination was blocked completely self-pollinated heterozygous ears would produce no Bf seedlings. If germination was not always affected or if it was just delayed, a less than expected number of Bf seedlings might be expected in some self progenies. 5) The mutation event could affect the viability of the female or male gametophyte. If the male gametophyte was involved, reduced transmission of the mutant allele would be expected in male outcrosses. If the female gametophyte was affected, reduced seed set would be expected.
Ten kernels from each of the outcrosses of 100 of the Bf1-Mu/Bf1-R heterozygotes were sown and the resulting plants (genotypes = Bf1-Mu/bf1 & Bf1-R/bf1) self-pollinated. In 38 of the outcross progenies tested, one or more of the progenies of the self-pollinated plants did not segregate for any Bf seedlings. To determine which of the 5 possible events described above was responsible for the absence of Bf seedlings, ten sibling kernels from each of these 38 families were sown and the resulting plants were screened for Bf anthers. This screening involved the systematic collection of the central spikes of all plants with freshly extruded anthers, arranged in a carrier that would allow the determination of the plant of origin for each. The anthers were screened in a dark room under ultraviolet light, and a washer was placed firmly on each spike with fluorescent anthers so that they could be recognized in normal light. All plants were screened for pollen sterility and, whether or not they had Bf anthers, they were reciprocally crossed to plants homozygous for the Bf1-R mutant allele. The seedlings from the progeny of the reciprocal crosses were then screened for the Bf phenotype (see Fig. 1).
In families of two of the 38 isolates, Bf1-Mu-044-3 and Bf1-Mu-544-3, all the plants lacked Bf anthers but some of the plants segregated for Bf seedlings in both the female and male outcross progenies to the homozygous Bf1-R tester stocks (Table 2). The Bf1-Mu-544-3 and the Bf1-Mu-044-3 isolates differed in that both female and male outcross progenies of all but 2 Bf1-Mu-544-3 plants (5033-5 & 7) segregated for 50% Bf seedlings, whereas most of the crosses of Bf1-Mu-044-3 had significantly less than 50% Bf seedlings in the progenies of both the male and female outcrosses. There was not a significant difference in the percentages of Bf seedlings in the female and male outcross progenies of this mutant (Table 2). These two isolates also differed in two other respects. While no outcross progenies of Bf1-Mu-044-3 had 100% Bf seedlings, two plants of Bf1-Mu-544-3 (5033-5 & 7) had only Bf seedlings in their outcross progenies and thus were homozygous for the mutant allele. These observations along with the lack of pollen abortion (see Tables 2 & 3) rule out deletions as an explanation for the lack of Bf seedlings in the original self progenies. The Bf1-Mu-544-3 isolate also differed from Bf1-Mu-044-3, in that the former isolate had two plants (5033-1 & 5) with an occasional anther with a small discrete fluorescent spot. Such spots were never observed in any plants of Bf1-Mu-044-3. The discreteness of these spots suggests that the anthranilic acid in this mutant is cell limited and not diffusible. Similar sharp boundaries between revertant nonfluorescent and mutant fluorescent sectors were found by Bender and Fink (Cell 83:725, 1995) in an analysis of the pai mutant of Arabidopsis, which accumulates anthranilate derivatives and exhibited blue fluorescence similar to Bf1-Mu-044-3 and Bf1-Mu-544-3.
To determine if these two isolates retained the same anther phenotype under the different environmental conditions of a second growing season and to ascertain the frequency of homozygous mutant seedlings, if they occurred, sibling kernels to those that produced the families tested in Table 2 were sown. In the family that produced the self-pollinated ear that was the source of the plants for tests of mutant Bf1-Mu-544-3 in Table 2, two sibling plants also had no Bf seedlings in the progenies of their self-pollinated ears. Plants from the kernels of the self-pollinated ears of these two plants were also included in these tests (Table 3). None of the tested families in Table 3 had any plants with totally fluorescent anthers. Plants 6 and 7 in family 1013 of mutant Bf1-Mu-544-3, however, had an occasional anther with a single small discrete fluorescent spot. The total Bf seedlings in the progenies of self-pollinated heterozygous plants of both isolates were significantly less than 25%. Although 11 out of the 31 progenies had a 3:1 ratio or a ratio close to 3:1, the numbers of Bf seedlings in most progenies were so few that the tests for significance were not very reliable. Thirteen of the progenies had significantly less than 25% Bf seedlings. Nine of these latter progenies had 2 or less Bf seedlings, including five that had no mutant seedlings. The lower than expected 25% mutant seedlings in the progenies of self-pollinated plants is not surprising because the ears chosen to provide the kernels for the plants involved in these tests did not segregate for any Bf seedlings. Although the original seedling tests that were made, by which the ears were chosen for further testing, consisted of about 50 seedlings, the results from the selfs in Table 3 suggest that the ears selected for the tests did carry some kernels that were homozygous for the Bf-Mu alleles.
The results from Tables 2 and 3 for Bf1-Mu-044-3 suggest that the lack or infrequent occurrence of Bf seedlings in progenies of selfs could be the result of kernels with homozygous mutant germs that did not germinate or germinated at less than the expected frequency. If homozygous mutant kernels did germinate, the Bf seedlings they produced did not go on to produce mature plants. This latter conclusion is confirmed by the observation that no mature plants were produced that were homozygous for the mutant allele in either Tables 2 or 3. Alternatively, the results from Table 2 suggest that the gametes with the mutant allele were transmitted in less than the expected frequency through both the male and female. Such transmission deficiencies also would result in less than the expected frequency of mutant seedlings in the progeny of self-pollinated plants. However, in the male outcrosses of this mutant in Table 3, 50% Bf seedlings were observed in about half of the seedling tests of the outcross progenies. The remaining plants had mutant percentages that approximated those observed for the male outcross in Table 2. Interestingly, however, most of the plants with 50% Bf seedlings in the male outcrosses, which had a sufficiently large number of seedlings in the self progenies for a good test of significance, had fewer than the expected mutant seedlings. These results could be due to a reduced transmission of the mutant allele because the female gametophytes with the mutant allele are lethal, or because the homozygous mutant zygote is lethal or because homozygous mutant germs do not germinate or, if they do, germination is delayed. Lethality of the female gametophyte and failure of homozygous mutant kernels to develop are ruled out because there is no consistent evidence of reduced seed set on self-pollinated ears (see Table 3). Zygotic lethality is not likely because it would result in germless kernels which were not consistently found to be associated with the transmission of these mutant alleles (see Table 3). Thus it would seem that reduced viability of homozygous mutant germs or the delayed germination of kernels with such germs is the likely explanation for the less-than-expected Bf seedlings in these self progenies.
Data from plants derived from the original self-pollinated ear of mutant Bf1-Mu-544-3 suggest that the allele is transmitted in expected frequencies when heterozygous plants are used in both male and female outcrosses (Table 2). This is also observed to be true in the male outcrosses of plants from sibling kernels of the same ear in Table 3 (families 1010 & 1011). However, heterozygous plants from sibling self-pollinated ears to the original one used in tests of this mutant (Table 2) exhibit a significant deficiency of mutant seedlings in male outcross progenies in the self and outcross tests of Table 3 (Families 1013 & 1014). The results from the latter two families are similar to those observed for the self and outcross tests of Bf1-Mu-044-3.
There is one outstanding difference between the results observed for these two mutants in these two sets of crosses (Tables 2 & 3). No homozygous Bf1-Mu plants are observed in the Bf1-Mu-044-3 families, while there are nine homozygous Bf1-Mu-544-3 plants in Tables 2 and 3. A total of 39 plants with this mutant allele were involved in the tests recorded in these two tables. Besides the nine mutant homozygotes, there were 12 plants homozygous for the nonmutant (bf1) allele and 18 heterozygotes. The ratio of these genotypes does not differ significantly from the 1:2:1 ratio expected if the mutant allele had normal transmission and if homozygous mutant plants were not lethal. If homozygous mutant plants were observed in these tests, why the lack of Bf seedlings in the progeny of the original self-pollinated ears and the deficiency of Bf seedlings in the progenies of self-pollinated plants derived from these original ears (Table 3)? Chance exclusion of Bf seedlings in the original seedling test of the self progenies of the three plants used in the test of this mutant seems very unlikely in light of the additional results from tests of sibling plants produced by kernels from the same ears (Tables 2 & 3). In the original seedling tests of this mutant, the seedlings were screened with ultraviolet light when most of the seedlings were at about the three leaf stage. If the homozygous Bf1-Mu-544-3 kernels were much slower germinating than the heterozygous or homozygous nonmutant kernels, Bf seedlings may not have emerged by the time these samples were screened, but would have perhaps emerged if the seedlings had been allowed to grow for a longer period of time before they were discarded. This also might explain the lower than expected percentages of Bf seedlings in the self progenies of all three populations of this mutant (Table 3, families 1010, 1011 - 13.40%, 1013 - 1.44% & 1014 - 12.12%). The seedlings screened in these latter tests came from the next generation of self-pollination and thus were not screened at the same time or under exactly the same conditions and perhaps had grown slightly longer before screening than the seedlings of the original self progenies, which may account for the presence of a few Bf seedlings in these latter tests (Table 3). If once the Bf seedlings emerge they are capable of developing into mature plants, the genotypes of mature homozygous nonmutant, heterozygous and homozygous mutant plants would be expected in the observed ratio.
The presence of a few Bf seedlings in the self progenies of Bf1-Mu-044-3 (Table 3) may also be the result of slow germination of the homozygous kernels. This mutant differs, however, from Bf1-Mu-544-3 in that homozygous Bf1-Mu-044-3 plants are not observed. Bf1-Mu-044-3 could be a more severe mutation, which when homozygous produces some Bf seedlings that die before the mutant plants can reach maturity.
The results from the tests of Bf1-Mu-544-3 differ from those of Bf1-Mu-044-3 in another way. Four Bf1-Mu-544-3 plants (Table 2 - 5033-1 & 5, Table 3 - 1013-6 & 7) had a unique anther phenotype. In the tassels of these plants, a few anthers were observed with a single small discrete Bf spot. These spots could possibly represent sectors of cells in which the Mutator element has transposed within the gene to a site that allows the mutant allele to be expressed in the tissue of the anther. Alternatively, the element may have transposed from the gene leaving a footprint that altered the expression of the mutant allele in such a way that it is expressed in the anthers as well as in the seedlings.
Of the 38 Bf1-Mu isolates that have been involved in this study only two lacked expression of the mutant phenotype in the anthers. Such a phenotype could result if these two alleles lack fluorescence in the anthers because they are insertion mutants in which the inserted Mu element is susceptible to suppression (modification) resulting from methylation of the inserted element. Martienssen, et al. (Genes Dev. 4:331, 1990) and Martienssen and Baron (Genetics 126:1157, 1994) describe a pattern of modification of Mutator-induced mutants that occurs progressively as the plant develops. Frequently the plants carrying the mutants they studied would have tassels in which the mutant phenotype was completely suppressed (nonmutant). If this phenomenon was responsible for the phenotype of these two Bf1-Mu mutants, it has so far been observed to occur in 100% of the heterozygous and homozygous mutant plants tested. Although over 1000 Bf seedlings were observed in these tests, not one had any sectoring in the later developing seedling leaves that might be expected if modification was occurring. Martienssen and Baron (1994) found that once a suppressible Mutator-induced mutant allele is modified it is not usually reactivated unless crossed to an active Mutator stock. Because the Bf1-R tester stocks used in the outcrosses in Tables 2 and 3 do not carry an active MuDSR element, no Bf seedlings would be expected in these outcrosses. However, only Bf seedlings were observed when homozygous Bf-Mu plants were outcrossed as males to Bf1-R tester plants. Likewise, the segregation of Bf seedlings when heterozygous plants were crossed to the same tester stocks is not expected if modification is occurring. These observations suggest that modification is not responsible for the unusual phenotype of the Bf1-Mu alleles. However, the possibility that some as yet not understood form of modification is responsible for the loss of the expression of blue fluorescence in the anthers of plants with these mutant alleles can not be eliminated as a possible explanation for their unexpected phenotypic expression.
These mutant stocks are potentially useful for the following kinds of studies: 1) Cloning the bf1 gene. 2) Studying the regulation of a gene that is expressed as a recessive in some tissues (seedlings) and as a dominant in other tissues (anthers). 3) The possible role of modification in the unique phenotype of these alleles.
Stocks of these and the other Bf1-Mu mutants are available upon request.
Table 1. Description of symbols used to designate alleles and phenotypes
involving the blue fluorescent 1 locus.
| bf1 | Symbol designation for this locusa |
| bf1 | Nonmutant allele at this locusa |
| Bf1-R | Mutant reference allele |
| Bf1-Mu | Any mutant allele induced by the Mutator system |
| Bf1-Mu-044-3 | Mutator-induced allele described in this note |
| Bf1-Mu-544-3 | Mutator-induced allele described in this note |
| Bf | Symbol used to designate the blue fluorescent phenotype as expressed in seedlings and(or) anthers of plants homozygous or heterozygous for a mutant allele |
| bf | Symbol used to designate the nonmutant phenotype |
Figure 1. Schematic diagram of the method used to produce and analyze the Mutator-induced Bf1-Mu mutants described in this study.
Table 2. Segregation of Bf seedlings in reciprocal crosses to homozygous Bf1-R plants of Bf1-Mu isolates that lacked Bf anthers.a
Table 3 A
& B.
Self and outcross tests of mutants Bf1-Mu-044-3 and Bf1-Mu-544-3,
which lack the Bf phenotype in the anthers but retain the seedling Bf phenotype.a
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}