Transcription has been shown to initiate at several sites for most maize mitochondrial genes. Transcription of the cox3 gene occurs at three sites which all possess the CRTA tetranucleotide sequence proximate to the transcription initiation nucleotide (Mulligan et al., Proc. Natl. Acad. Sci. USA 85:7998, 1988). Two of these three sites (at -360 and -320 relative to the ATG) show extensive homology to each other over a 21 nucleotide stretch; this repeated sequence can therefore be defined as a minimal promoter sequence. The close proximity of the two promoters allowed for the simple construction of a probe for use in mobility shift assays to identify the proteins involved in plant mitochondrial transcription.
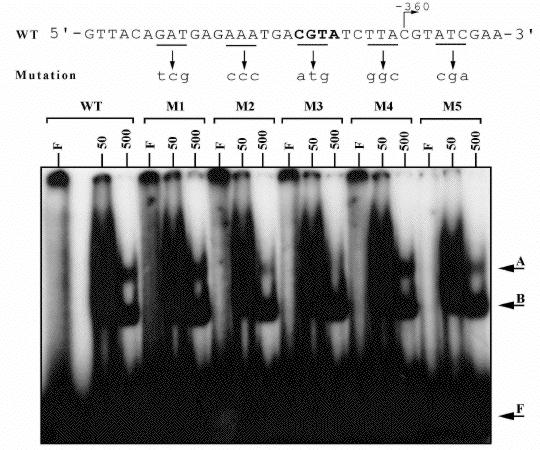
Initial studies with the double-promoter probe and a S-100 mitochondrial protein extract produced a complex retardation pattern that was presumed to be due to the binding of a number of proteins to both promoter elements. After dividing the probe into its constituent promoter elements, -360 and -320, and using each promoter element individually in mobility shift assays the retardation pattern was simplified. The retardation patterns of each promoter were identical displaying two slowly migrating complexes, named A and B. These retardation complexes were stable in high-salt, 500 mM KCl, and in the presence of a 2000-fold excess of non-specific competitor DNA demonstrating the specificity of the interactions. Linker-scanning mutagenesis of the cox3-360 promoter identified the sequence CRT of the 5'-CRTA-3' motif as being essential for complex-A formation (Fig. 1). Furthermore sequences both upstream and downstream of the CRTA motif were shown to be required for optimal complex-A formation (Fig. 1).
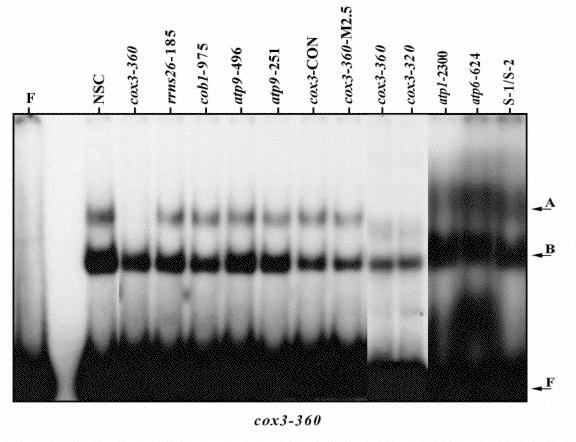
Mobility shift competition studies were performed to address the question as to whether complex-A formation was specific to cox3 or whether the binding factor could recognise other mitochondrial promoter sequences. The principle of these experiments was to add an excess of unlabelled specific competitor DNA to a reaction containing the radiolabelled cox3-360 probe and the S-100 mitochondrial protein extract. If the competitor DNA was capable of sequestering the complex-A binding factor then a reduction in the radiolabelled complex-A would be observed. Figure 2 shows the result of such an experiment. When the competitor DNA added to the reaction was either of the cox3 promoter sequences (-360 or -320) the formation of complex-A was greatly reduced due to the sequestration of factor `A` by these DNAs. However when the competitor DNA used was another characterised maize mitochondrial promoter sequence (see figure legend for details) complex-A could still be visualised. These results therefore indicate that factor `A` binds specifically to the cox3 promoter elements at -320 and -360. This result is surprising and strongly suggests that mitochondrial transcription requires `promoter-specific` factors. A permutation of different promoter sequences upstream of genes will reduce the reliance of gene transcription on the abundance of any one factor. The recent proposal of gene-specific transcription of the cox2 gene in Zea perennis associated with the MCT locus (Newton et al., EMBO J. 14:585, 1995) supports this theory that each plant mitochondrial gene may require its own factor for accurate transcription initiation.
Figure 1. Mobility shift experiment using mutant cox3-360 probes. The sequence of the cox3-360 `wild type' probe is shown above the autoradiograph. On the sequence the nucleotide at which transcription initiation occurs is indicated with an arrow. Underlined are the triplet sequences that were mutated to form the mutant probes. The sequences of these mutated probes (M1 to M5) are shown in lower case above their corresponding lanes. Each probe was labelled with [ÿ-32P] dATP and [ÿ-32P] dTTP, and 12,000 cpm of the probe were loaded in each of three lanes. The first of the three lanes F contained only probe, the second and third (50 and 500 respectively) contained 15 µg S100-mitochondrial protein and the labelled quantity (in ng) of non-specific competitor DNA (polydIdC.polydIdC). The arrows labelled A, B and F refer to bound complex-A, complex-B and free probe respectively.
Figure 2. Competition study using various `cold' mitochondrial promoter fragments versus radiolabelled cox3-360. All lanes contained 12,000 cpm of the cox3-360 probe and all, except lane F (free probe), contained 15 µg S100-mitochondrial protein and 500 ng polydIdC.polydIdC. Unlabelled competitor double-stranded oligonucletides, named above their corresponding lanes, encompass other maize mitochondrial promoter sequences, except where noted below, and were present in the reactions in a 50-fold excess relative to the labelled probe. NSC, no specific-competitor; cob1-975 is a rice mitochondrial promoter; cox3-CON is a 5' flanking region of the maize cox3 gene that contains a CRTA motif yet does not sequester factor `A`; cox3-360-M2.5 is a TGA-mutation between mutations M2 and M3 (see Fig. 1) and S-1/S-2 is the promoter present in the terminal inverted repeat of the maize cms-S plasmids.
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}