Analysis of nitrogen partitioning in maize
--Balconi, C; Bosio, D; Motto, M
Efficient use of nitrogen (N) fertilizer for maize production is important to maximize economic return to the grower, to minimize the potential impact on water quality, and to reduce the total energy required for manufacture of N. Current work indicates that the efficiency with which maize plants utilize N fertilizer is affected by several parameters including root morphology and extension, biochemical or physiological mechanisms regulating NO3 uptake, redistribution and transport of N to different plant parts (Rizzi et al., Maydica 40:253-258, 1995). Therefore, the potential for developing superior N-efficient hybrids appears to exist. Evidence shows that maize can absorb substantial quantities of N following anthesis, and mobilization of vegetative N accumulated before anthesis provides the major source of N in the grain. It has been observed that N in the stalk of maize during vegetative growth can constitute as much as 40-50% the total-N content of the plant (Ta, Plant Physiol 97:1375-1380, 1991). Clearly, at least in some circumstances, the stalk is an important temporary reservoir for N that can be remobilized for ear development.
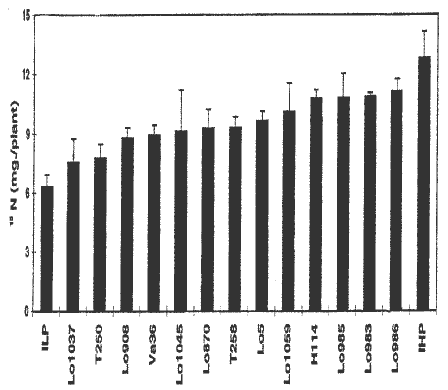
In this investigation we have analyzed 13 inbred lines of maize, previously identified with high or low N-NO3 stalk content at anthesis, and the Illinois High Protein (IHP) and Illinois Low Protein (ILP) strains, previously tested (Bosio et al., MNL 69: 19-20, 1995). Plants were grown under field conditions at Bergamo, during 1995, to document the partitioning of 15N applied to the different genotypes, and to follow the pattern of subsequent remobilization of this N during ear development. For each genotype 340 mg of labeled N (as K15N03, 14.4 atom % excess, corresponding to 50 mg N labeled) were applied to individual plants in the two central rows of each subplot. In Figure 1 the 15N total plant content at black layer maturity (BLM) is reported for the genotypes tested. It is evident that substantial phenotypic variation in the N-uptake efficiency exists.
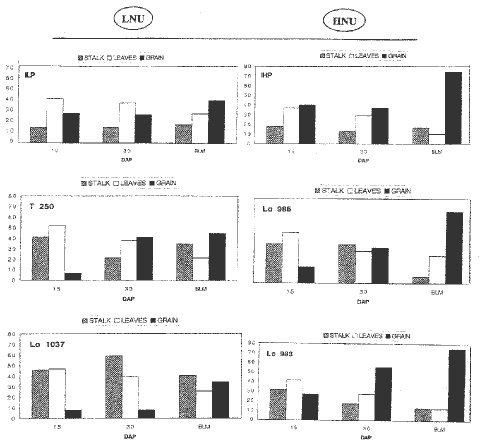
For each subplot plant samples were harvested at 3 stages during grain filling: 15 and 30 days after pollination (DAP) and at BLM. The plants were divided into stalks, leaves and kernels for 15N determination. In Figure 2 the distribution of 15N in plant parts is reported for the genotypes identified for low (LNU: ILP,T250,Lo1037) and high (HNU: IHP,Lo986,Lo983) N-uptake. In the HNU plants we observed a higher relocation of 15N in the kernels than in the LNU plants; on the other hand, in the LNU plants 15N content in the stalk at BLM was higher than in HNU plants. These data suggest that the stalk plays an important role in storing and providing N for kernel development. It is proposed that this trait could be used as a selection criterion for developing more efficient plants in N utilization.
Figure 1. Plant 15N content (mg/plant) at BLM in the genotypes tested. Vertical bars indicate standard errors.
Figure 2. Distribution of 15N in plant parts in the genotypes identified for LNU (ILP, T250, Lo1037) and HNU (IHP, Lo986, Lo983).
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}