Visualization of methylated cytosine in maize somatic chromosomes
--Fluminhan, A; Ohmido, N; Fukui, K; Mizugaki, M; Kameya, T
Methylation of DNA is considered to be a determinant of gene expression and its role in growth and development has been extensively studied in many systems. Diverse evidences have implicated the changes in the pattern and degree of DNA methylation in the control of a number of cellular processes, including gene activity, cellular differentiation, chromatin structure, DNA replication, transcription and repair. Methylation interferes with DNA-protein interactions and thus may play a crucial role in all processes involving such interactions. However, much remains to be elucidated about this role and about the distribution of methylated bases at the DNA and chromosome levels. The DNA of higher plants contains a considerable proportion of cytosine residues modified to 5-methylcytosine (5-MeC). Up to 30% of the cytosine residues are methylated in certain plant species, compared to 3 to 8% in vertebrates (Adams and Burdon, Molecular Biology of DNA Methylation, Springer-Verlag, 1985). Virtually, all of the 5-MeC found in plant genomes is confined to the symmetrical sequences CpG and CpNpG, which are distributed non-randomly in the DNA. Many studies have demonstrated that certain genes are undermethylated in tissues where they are actively expressed, but highly methylated in tissues where they are silent, suggesting that methylation of DNA is a possible mechanism by which gene activity is regulated during development. The relationship between the degree of DNA methylation and gene expression has been found primarily in transgenic plants (Amasino et al., MGG 197: 437-446, 1984) and transposon insertions (Chandler and Walbot, PNAS 83:1767-1771, 1986 Schwartz and Dennis, MGG 205:476-482, 1986).
Immunological detection of 5-MeC has been used in studies with mammalian cells to detect different levels of methylation between euchromatin and heterochromatin, and between the heterochromatic regions of different chromosomes in the same complement (Miller et al., Nature 251:636-637, 1974 Schnedl et al., Chromosoma 59:59-66, 1975). High levels of 5-MeC are found in unexpressed regions of human and mouse genomes, such as centromeric and heterochromatic regions, several types of satellite DNA, and inactive mammalian X-chromosomes. We describe here an indirect immunofluorescence procedure based on the use of anti-5-MeC antisera for the analysis of the distribution of methylated cytosine in somatic chromosomes of maize. Chromosome preparations from long-term cultured cells and root meristems of germinating aged seeds were employed. Single-stranded DNA was produced either by UV irradiation or by heating in a formamide solution. In the first case, slides were placed cell side up in a plastic Petri dish and covered with a layer about 1 cm deep of PBS solution (phosphate-buffered saline: 20g NaCl, 85 ml 0.25M Na2HPO4, 15 ml 0.25M KH2PO4 in 2400 ml distilled water, pH 7.2 - 7.4). The dish was placed 32 cm from two 15 W germicidal lamps (Hitachi) and exposed to UV irradiation for 18 h at room temperature. For the heating treatment, chromosome preparations were incubated in a 90% formamide solution with 0.25% formaldehyde for 60 min at 65 C. Slides were then covered with an appropriate dilution of specific anti-5-MeC antisera (elicited in mouse by immunization with nucleoside-protein conjugates) and incubated in a moist chamber for 30 min at 37 C. The preparations were rinsed with PBS solution, layered with an appropriate dilution of specifically purified goat anti-mouse immunoglobin tagged with biotin and incubated for additional 30 min at 37 C. After staining with fluorescein-isothiocyanate (FITC)-avidin conjugate, signals were amplified by applying a biotinylated anti-avidin solution to the preparations, followed by incubation with a fluorescein-avidin solution. Chromosomes were then counter-stained with DAPI, and examined by fluorescence microscopy. Images were captured by a cooled CCD camera (Photometrics) mounted on the microscope. Digitized images were photographed by a color image recorder (Nippon Avionics).
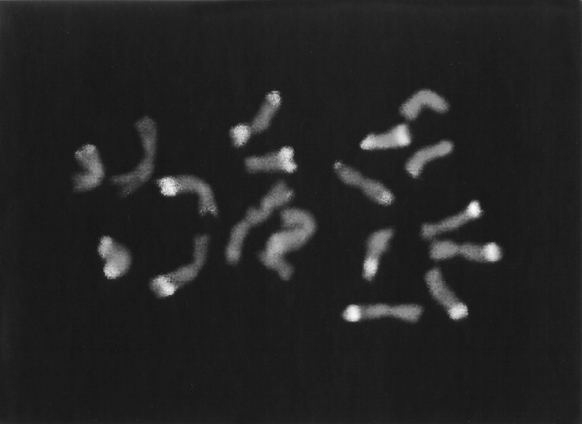
Our results are consistent with the binding of anti-5-MeC antisera on single-stranded DNA in fixed chromosomes. The antibody employed in the present study was observed to bind intensely to specific regions in maize somatic chromosomes, reflecting a non-random distribution of 5-methylcytosine in the nuclear DNA of this species. Brilliant fluorescence, indicating extensive antibody binding, was mostly observed in knob heterochromatic regions (Figure 1). The degree of fluorescence and the size of the fluorescent regions tended to vary in different cells and in the two homologues of the same complement. The intense fluorescence of heterochromatic knobs partly reflects the greater concentration of 5-MeC at the knob repetitive sequences. We have also observed a moderately bright fluorescence in the distal portion of several chromosome arms, and in the nucleolus-organizing region. The fluorescent pattern observed with anti-5-MeC antisera might reveal the possible location of CpG and CpNpG-rich regions of the maize genome. Consistently, Dennis and Peacock (J. Mol. Evol. 20:341-350, 1984) have reported that the 180 bp knob-specific repeats could show up to ten sites with CpG dinucleotides and/or CpNpG trinucleotides among the different clones sequenced. It is recognized that the binding pattern of the anti-5-MeC antibody might also reflect differences in regional base composition, extent of denaturation of DNA, density of nucleoproteins, tertiary structure or protein binding. Furthermore, current evidence suggests that 5-methylcytosine is not connected with maintenance of the stability of eukaryote DNA or DNA synthesis. Thus, DNA from different tissues of the same plant (or different developmental stages of the same tissue) may have a differing content of 5-MeC.
The observation of an increased presence of methylated cytosine at knob heterochromatin is consistent with the proposition that changes in the degree and pattern of DNA methylation could be an underlying cause of chromosomal abnormalities commonly observed in maize tissue cultures and their regenerants (Phillips et al., Proc. 7th Intl. Cong. Plant Tissue Cell Cult. pp. 131-141, 1990). These findings have prompted us to investigate the role of this biological modification in chromosomal abnormalities currently observed in cultured cells and other systems, such as for example: a) at first mitosis in germinating aged seeds (Fluminhan and Kameya, Genome - in press) and b) at the second microspore mitosis in a specific genetic background where the combination of B chromosomes and knobs leads to chromatin loss (Rhoades et al., PNAS 57:1626-1632, 1967; Rhoades and Dempsey, Genetics 71:73-96, 1972; Rhoades and Dempsey, J. Hered. 64:12-18, 1973).
We can conclude that indirect immunofluorescence procedures should therefore be very useful in analyzing the nature of the DNA present in the chromosomes, especially because of the high degree of base specificity shown by antinucleoside antibodies. Our results clearly show the applicability of the method for the direct visualization of methylated cytosine in maize chromosomes and may represent a promising approach to the molecular structure of nuclear genomes and the evolution of highly repetitive sequences.
Figure
1. Metaphase chromosomes of a 2-year-old maize embryogenic callus culture
after indirect immunofluorescence detection of 5-methylcytosine (5-MeC).
Higher levels of methylated cytosine are observed at the heterochromatic
knobs.
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}