Localization of autonomous pairing sites in maize
--Goldman, SL; Doyle, GG; Blakey, CA
The existence of autonomous pairing sequences (APS) which function in the process of homologous recognition have been postulated by many scientists working to define the mechanism of homologue sorting. Using the convention established by Sybenga (Genetica 37:186, 1966), the term zygomere is used to describe those genetic elements, i.e., nucleotide sequences, specialized to function in the initial phases of pairing. Given this, we are identifying the segments associated with zygomere activity using the technique of Restriction Fragment Length Polymorphism (RFLP) mapping. This is made possible by the fact that trisomic stocks segregating heterologous zygomeres have been mapped to chromosome 3 (Doyle, TAG 79:145, 1990). These autonomous pairing sites have been recognized following changes in preferential pairing and should associate with concomitant alterations in RFLP profiles subsequent to Southern analysis. As a result, an unambiguous linkage should exist between the phenomenon of preferential pairing and the segregation of heterologous zygomeres. In principle, this association is detected when seed lines exhibiting marker segregation distortion co-segregate with (a) defined RFLP(s).
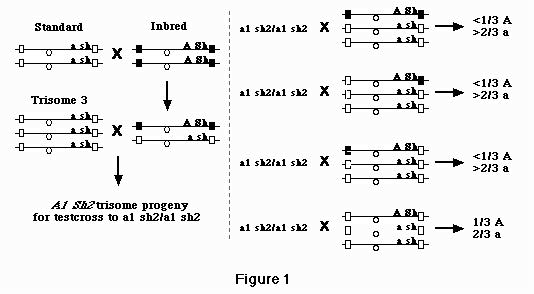
The inbreds B41, Hy, and 38-11 have been shown to be marked with distinctive APSs. Therefore, if the F1 hybrid (standard a1 sh2/inbred A1 Sh2), for example, is crossed to the standard trisome (a1 sh2/a1 sh2/a1 sh2), A1 and Sh2 will segregate from their respective APSs in the progeny. This conclusion is based on the fact that at least one autonomous pairing site and A1 Sh2 region should recombine freely if these synaptic initiator sequences map to the subterminal ends of chromosome 3 as has already been shown. As such, it will be possible to recover chromosomes marked by A1 Sh2 whose pairing kinetics have changed. The location of putative autonomous pairing sites on chromosome 3 has been recognized following changes in preferential pairing, and they associate with concomitant alterations in RFLP profiles subsequent to Southern analysis. We are in the process of analyzing the segregation pattern of each of the 10 RFLP core markers of chromosome 3 in lines segregating preferential pairing.
Plants of the inbred lines (B41, Hy, and 38-11) have been crossed with the multiple marked stock noted above. The hybrids obtained will be heterozygous for all gene loci and crossed as the pollen parent with standard trisome 3 which is homozygous for al and sh2 and the other six dominant alleles. These trisome 3 heterozygous (A1 Sh2/a1 sh2/a1 sh2) are crossed as the pollen parent onto a1 sh2 testers to determine the presence or absence of preferential pairing as indicated by changes in the ratios of A1 Sh2 to a1 sh2 in the progenies. In addition, each tested trisome is self-fertilized to determine which chromosome was donated by the diploid hybrid parent. In this way, linkages between preferential pairing factors (heterologous zygomeres) and genetic markers can be established. Following the mapping, we will expand these lines in order to obtain greater quantities of fresh material. For the initial study only A1 Sh2 markers will be used as illustrated in Figure 1.
Figure 1 outlines a simplified series of crosses designed to illustrate the correspondence between zygomere activity and preferential pairing. The black and white squares represent zygomeres in different mutational states. For the purpose of discussion their placement is arbitrary. The only difference in the four test crosses represented is expected to be a function of heterologous zygomere expression. This will be determined by showing a correspondence between seed having modified genetic ratios and (a) defined RFLP(s). Specifically, plants from test crosses 1, 2 and 3 that give preferential pairing should segregate different RFLPs than those found in test cross 4 where pairing is at random. The diploid standard/inbred hybrid was also crossed to a diploid tester (a1/a1 or sh2/sh2) as a control to rule out the presence of gametophyte factors, differential viability of gametes or zygotes, or anything that would disturb the genetic ratios. Here, as expected 50% transmission of A1 Sh2 was observed.
DNA from the inbred lines and the standard stocks segregating the chromosome 3 markers as well as the resulting F1s have been restricted with EcoR1, HindIII and EcoRV. As a first screen, this DNA was probed, in turn, with RFLP markers mapping to the subterminal ends of linkage group 3. These enzymes have been proven to be particularly effective in elucidating polymorphisms in maize in the past (Hoisington and Coe, Gene Manipulation and Plant Improvement, p. 331-352, 1990). This simple test has discriminated the RFLPs of interest as they exist in this region of chromosome 3. Similar results have been obtained with the RFLP markers that span the length of chromosome 3.
Significantly, the material segregating the different zygomeres is highly polymorphic based on experiments using three restriction enzymes that include EcoRV, EcoRI, and BamHI and 10 different RFLP makers. Given this, the morphs that are associated with each inbred on a marker by marker basis can be delineated from those segregating in the standard. RFLP markers have been used to screen DNA isolated from inbreds, from standard/inbred hybrids, from trisomes, and from tester lines. Significantly, leaves from approximately 800 plants have already been isolated from the progeny of stocks that express preferential pairing and from the progeny of stocks that do not based on the transmission of A1 Sh2. In addition, DNA has already been isolated from a limited number test cross progeny using the cross standard zygomere/standard zygomere X B41 zygomere/standard zygomere/standard zygomere.
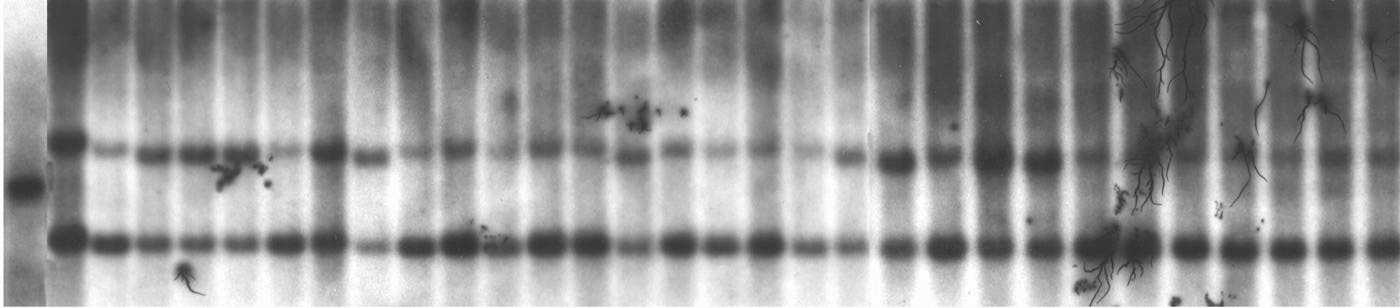
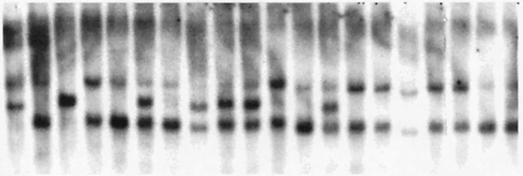
DNA from 67 plants representing three different families where preferential pairing is expressed based on seed marker segregation has been isolated, cut with EcoRV and probed for umc32. This marker is linked both to the proximal subtelomeric region of chromosome 3 (umc32a) and to the centric region of chromosome 8 (umc32b). When DNA from the inbred B41 is cut with EcoRV a single band is discriminated as is shown in lane 1 of Figure 2. When DNA is isolated from tester-lines segregating standard zygomeres and probed with umc32 two bands are distinguished as is shown in lane 2. Lanes 3 through 32 show the segregation pattern of B41:standard when DNA is isolated from individual plants obtained from a single ear. In this connection, B41 segregates from standard in a ratio of 0:29. Specifically, the B41 allele is not transmitted and evidence of RFLP segregation is absent.
When data from these 67 plants are pooled, the segregation pattern of B41:standard is also of interest. Specifically, B41 segregates from standard in the ratio 10 to 57. This distribution differs significantly from the expected 1 B41:2 standard when preferential pairing is absent. Differences in degree of preferential pairing could reflect the number of different heterologous zygomeres segregating in a population. Under the assumption that there is a minimum of one zygomere in each chromosome arm, three different populations of individuals expressing distinctive patterns of preferential segregation are possible as is shown in Figure 1.
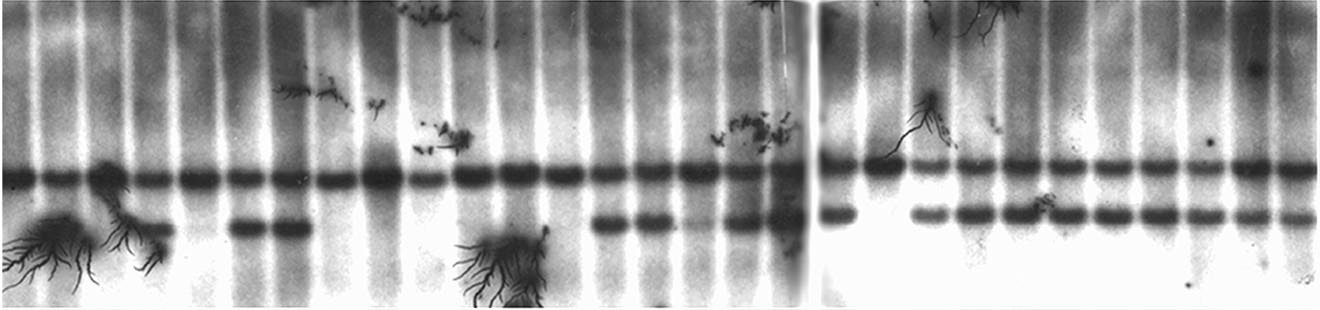
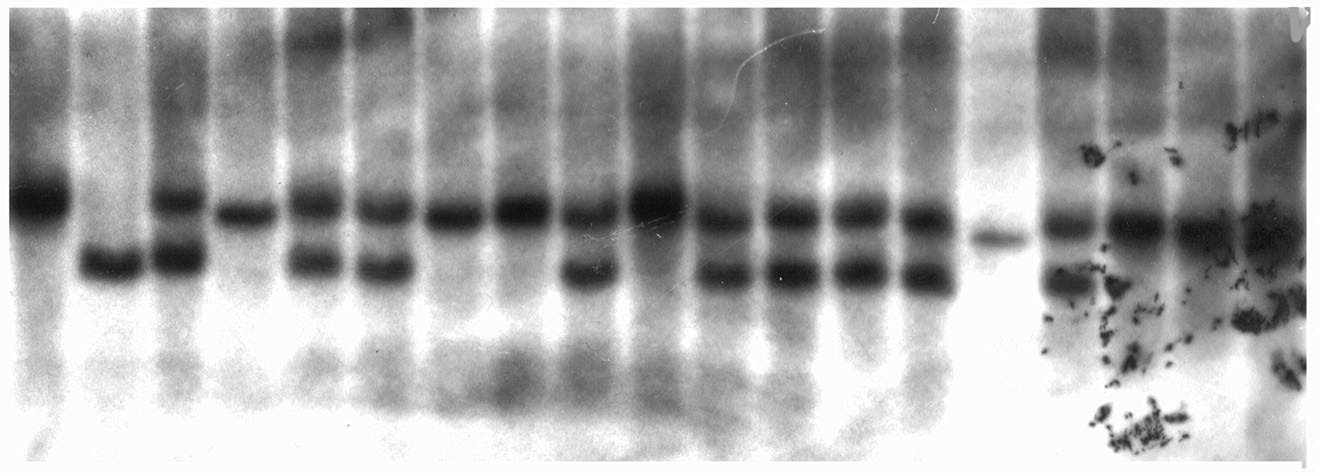
DNA from these same 67 plants was probed using the marker umc161. umc161 maps to the centric region of chromosome 3. A portion of this data is shown in Figure 3 and corresponds precisely to the plant DNA probed in Figure 2. When B41 is cut with EcoRI and probed with umc161 a single band is discriminated as is shown in lane 1. When the tester lines are probed a single distinctive signal is likewise identified. B41 segregates from standard in a ratio of 18:11 in this family. The presence of both morphs provides evidence that this family constitutes a segregating population. This observation is particularly important, since RFLP segregation was absent when this DNA was probed with umc32. Among the 67 plants scored B41 segregates from standard in a 36:31 ratio.
DNA has also been isolated from plants where preferential pairing is absent. When B41 is cut with EcoRV and probed with umc32, a single band is discriminated as is shown in lane 2 of Figure 4. When the tester lines segregating standard zygomeres are cut with EcoRV two bands are identified as is shown in lane 1. Lanes 3 through 20 are the segregants of the RFLP variants from the cross a1 sh2 (standard)/a1 sh2 (standard) X A1 Sh2 (B41)/ a1 sh2 (standard)/a1 sh2 (standard). In this connection the B41 morph segregates from standard in a ratio of 7 to 10. When this blot is reprobed with umc161 the morphs segregate in a similar manner, see Figure 5. Based on these most preliminary findings the segregation distortion associated with preferential pairing appears to be region specific and linked to the subtelomeric region of chromosome 3. These data lend credence to the hypothesis that if the zygomeres linked to chromosome 3 are marked by an RFLP, it should be possible to map these structures. Specifically, it should be possible to identify the RFLP(s) that co-segregates with preferential pairing and thereby establish the map position of the zygomere.
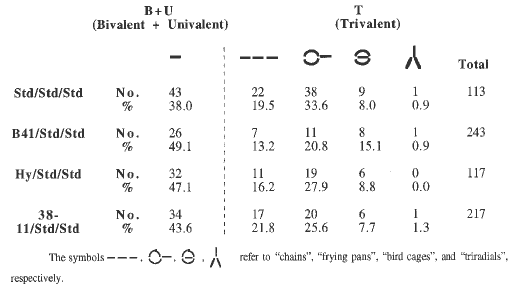
Work is in progress to demonstrate the expression of zygomere activity cytologically. In principle, trisomes manifest two general pairing modes that include either a bivalent and a univalent or a trivalent. Given this, four types of distinctive trivalents have been discriminated. These include "chains", "frying pans", "triradials", and "birdcages". Since preferential pairing is associated with the inbreds B41, Hy, and 38-11 but not with W23, there should be a reduction in the frequency of trivalents and shifts in the relative frequencies of each of the four types of trivalent. As predicted, the frequency of bivalents plus univalents rises from 38% in the control to 43%, to 49%, and to 47% in 38-11, B41, and Hy respectively. For details see Table 1.
From a study of chromosome pairing in the plant kingdom, it may be concluded that a complex of different mechanisms may function to delineate homologous recognition. For example, the prescriptive global response observed in allohexaploid wheat restricts recognition to intragenomic homologues in the presence of the Ph gene (Sears and Okamoto;, Genetics 2:258, 1958 Riley et al., J Hered 52:22, 1961). When this gene is deleted, the pairing kinetics change and homeologous associations form leading to a rise in multivalent formation and a concomitant decrease in the frequency of fertile gametes. Formally then, in the absence of Ph this alloploid is changed to an autoploid with attendant problems associated with fertility.
Given this, the RFLP mapping of zygomeres in maize will have enormous practical consequences for plant breeders. For example through the manipulation of zygomeres it may prove possible to turn a maize autotetraploid into an allotetraploid. Autotetraploids segregate a high frequency of aneuploids, which decrease the fitness of the populations (Doyle, TAG 54:103, 1979a; TAG 54:161, 1979b; TAG 61:81, 1982; TAG 71:585, 1986). Strictly speaking, an allotetraploid is a true breeding hybrid and would have few aneuploids. This said, considerable progress has already been made on the allotetraploidization of maize through the mutation of zygomeres (Doyle, 1979a, b; 1982; 1986; 1990). Specifically, the sequestering of homeologous chromosomes one from the other in these lines can not be attributed to chromosome aberrations as no cytogenetic anomalies have been observed. When the zygomeres are identified and mapped, restructured lines using conventional breeding protocols, could be synthesized with less difficulty as has been suggested by Sybenga (Euphytica 22:433, 1973), leading to the production of maize allotetraploids.
Table 1. Effect of preferential pairing on pairing configurations in trisomes.
Fig.
5
Please Note: Notes submitted to the Maize Genetics
Cooperation Newsletter may be cited only with consent of the authors
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}