Physical location of maize (Zea mays L.) cdc2 and prh1
genes by in situ hybridization
--Ren, N; Song, YC; Bi, XZ; Ding, Y; Liu, LH
A biotin-labeling in situ hybridization technique was used to physically map two single copy genes, cdc2 and prh1 in maize. The product of the cdc2 gene is the p34cdc2 protein kinase (ser/thr kinase), which is believed to be a central component controlling cell division in eukaryotes (Colasanti et al., Proc Natl Acad Sci USA 88:3377-3381, 1991). This protein kinase is a part of the M-phase promoting factor (MPF), which is the key regulation factor of the cell cycle (Nurse, Nature 344:503-507, 1990). The product of the gene prh1 is ser/thr phosphatase (type 1 protein phosphatase, PP1). It is speculated that plant PP1 could be involved in the regulation of a number of cellular processes including mitosis, chromosome separation, transcription, and protein synthesis (Smith et al., Plant Physiol 97:677-683, 19910. The reversible phosphorylation of proteins is a major mechanism by which metabolic and developmental processes are regulated in eukaryotic cells (Cohen et al., J Biol Chem 264:21435-21438, 1989). Therefore, these two genes are metabolically interrelated.
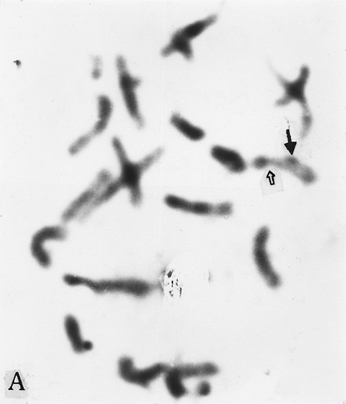
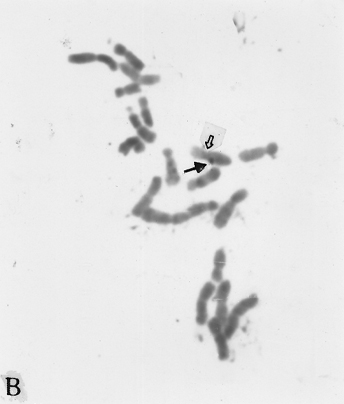
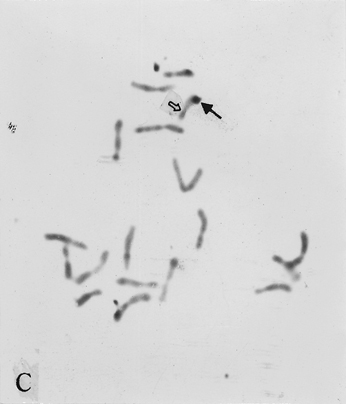
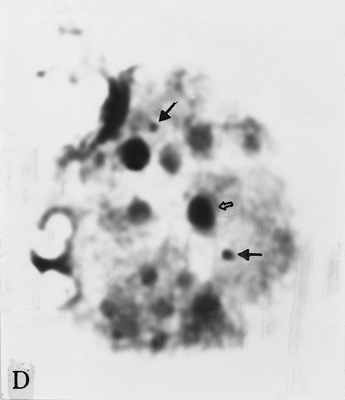
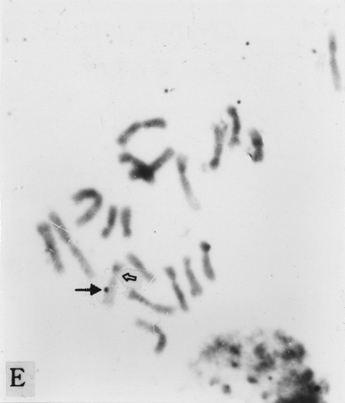
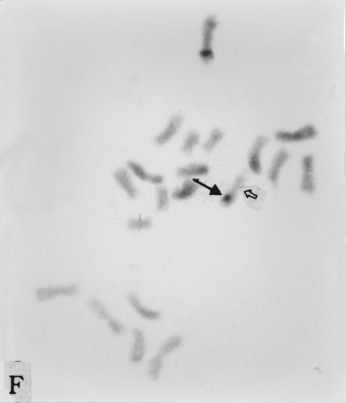
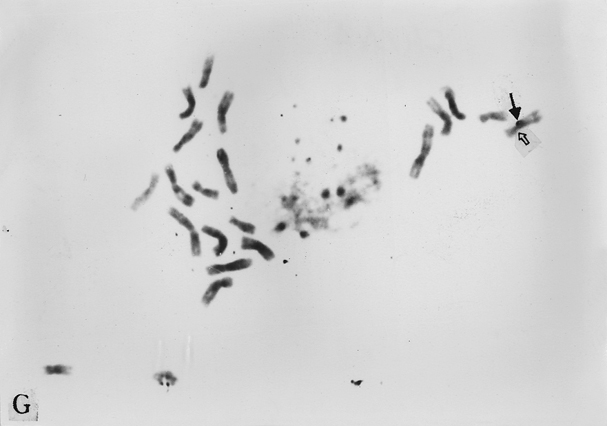
The full-length cDNA clones cdc2 ZmA and ZmPP1 of genes cdc2 and prh1 were adopted as the probes. They are 1.3 and 1.7 kb in size respectively. Clone cdc2 ZmA was physically mapped on the long arms of chromosomes 4, 8, and 9 (Fig. 1 A, B, C). The percent distances from centromere to detection site were 57.87 ± 2.68, 28.42 ± 1.45 and 88.16 ± 3.26 (Table 1). The detection rates were 10.07%, 3.13% and 8.33% respectively. Clone ZmPP1 was physically mapped on the long arms of chromosomes 4, 6 and 8 (Fig. 1 E, F, H). The percent distances were 53.62 ± 1.17, 60.77 ± 2.90 and 17.10 ± 1.61. The detection ratios were 12.07%, 5.17% and 6.17% respectively (Table 1).
For the hybridization of the cdc2 ZmA probe there were differences in detection rates among the different chromosomes. We think there must be differences in homology between the probe and the DNA sequence among different hybridization sites. The greater the degree of homology, the greater the chance that the probes were hybridized onto the chromosomes, and the more tightly the probes combine with the chromosomes. This suggests that the gene cdc2 is located on chromosomes 4 and 9, and that there is a gene for another protein kinase that has homology with the gene cdc2.
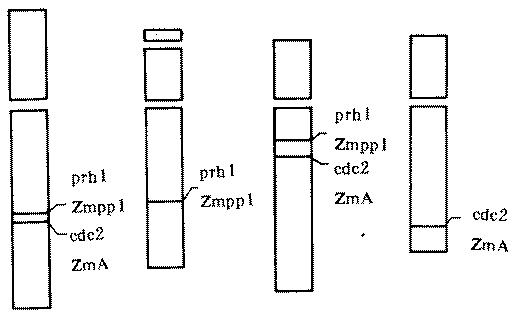
The facts that not only the genes cdc2 and prh1 are metabolically interrelated genes, but also that their physical locations are on chromosome 4 and near each other (Fig. 2), lead us to think about the relationship between the location and function of genes on a chromosome. As we know, the homeotic genes of Drosophila in a cluster on a chromosome are arranged in the order in which they are expressed along the body axis (Harding et al., Science 229:1236-1242, 1985; Akam, Development 101:1-22, 1987) and it has been proved by Blumenthal et al. that there may also be operon structure in eukaryotes. They reported that the nematode, Caenorhabditis elegans, adopted both the prokaryotic and the eukaryotic patterns of gene organization and transcription and at least a quarter of the genes seem to be organized into operons (Zorio et al., Nature 372:270-272, 1994). Whether the fact that the genes cdc2 and prh1 are linked closely represents the general trend that metabolically interrelated genes in eukaryotes, including plants, have an operon structure or an organization that resembles operons or not, is a significant problem which remains to be studied further.
Table 1. Average arm ratios and signal location in the chromosome in
the maize genome.
| Probe | Chr | Detections (No.) | Ratio of long to short arm | Signal location (%) |
| cdc2 | 4 | 29 | 1.72±0.05 | 57.87±2.68 |
| 8 | 9 | 2.81 ± 0.07 | 28.42±1.45 | |
| 9 | 24 | 2.01±0.05 | 88.16±3.26 | |
| prh1 | 4 | 28 | 1.80±0.07 | 53.62±1.17 |
| 6 | 12 | 2.70±0.16 | 60.77±2.90 | |
| 8 | 15 | 2.75±0.10 | 17.10±1.61 |
Figure 1. The hybridized sites of the probes cdc2 ZmA and ZmPP1. The black arrows show the hybridization signals in all figures, the white arrows show the chromocenters in d and h, and centromeres in the other figures. (A) The hybridization signal of cdc2 ZmA is in the long arm of chromosome 4. (B) The hybridization signal of cdc2 ZmA is in the long arm of chromosome 8. (C) The hybridization signal of cdc2 ZmA is in the long arm of chromosome 9. (D) The interphase cell has two hybridization signals of cdc2 ZmA. (E) The hybridization signal of ZmPP1 is in the long arm of chromosome 4. (F) The hybridization signal of ZmPP1 is in the long arm of chromosome 6. (G) The hybridization signal of ZmPP1 is in the long arm of chromosome 8. (H) The interphase cell has five hybridization signals of ZmPP1.
Figure 2. Ideograms of hybridization sites of cdc2 ZmA and prh1 ZmPP1 probes.
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}