The theoretical significance of a mutant in the phylogeny of the
species Zea mays L.
--Radu, A; Urechean, V; Naidin, C; Motorga, V
"The whole living world has had one single start point. Theoretically speaking, if we would have studied this point, it is possible to find out several genes that have been kept just as they were for three billion years." (Maximilian C., Doina Maria Ioan, Dict Enc Genet, Stiint Enc Bucharest, 1984)
The origin of corn is still submitted to scientific controversy because of the morphological changes that led to the appearance of the corn plant as it is known today, deriving from the ancestral, wild extinct forms (Galinat, pp. 1-4 in Sprague, Corn and Corn Improvement, Amer Soc Agron, Madison, 1977). The Maydeae tribe includes seven genera: two of American origin, Zea and Tripsacum, and another five of Eastern origin, Coix, Chionachne, Sclerachne, Trilobachne and Polytoca. It is generally agreed that the greatest role was played by the American genera Zea and Tripsacum in corn phylogeny, whilst the Eastern ones played only a small role. It is accepted that the Coix genus might have occurred in a certain stage of the phylogenetic development of the species Zea mays L. There are a number of characters that show phenotypic expression and penetration according to which we presume that the Manisuris genus, belonging to the Andropogoneae, might have contributed to the phylogenetic tree of final development of Zea mays L.
Cytogenetic studies at the Zea x Tripsacum hybrid level (Mangelsdorf and Reeves, Adv Genet 1947) confirmed the presence of several Tripsacum chromosomes, on which the supernumerary B chromosome existence relies and that can be found in the current corn genotypes (Galinat, 1977). Undoubtedly, the origin of Zea mays relies on an ancestral extinct corn form with genome (2n = 20 MM) and many recessive genes that led to the appearance of a new sterile hybrid F1 (2n = 10 M + 18 T) by natural hybridization to Tripsacum dactyloides (2n = 36 TT).
By natural diploidization a new hybrid form F1 (2n = 20 MM + 18 T) has been obtained, which led to the appearance of the old corn forms by introgressive hybridization to the ancestral corn form (2n = 20 MM) under the influence of empirical selection. This model of hypothetical development has been checked and confirmed (Galinat, 1977), but using as a starting point old American corn varieties. The ancestral corn form that initiated the phylogenetic development of Zea mays by natural introgression is unknown today. The various natural mutants that appear with Zea mays do not show special scientific interest as far as the origin of corn is concerned, the mutant gene expression being within the limits of intraspecific variation. Up to now (February 1996) there were no reports about corn mutants that might explain the development of the Zea genes by the contribution of certain species belonging to genera outside the Maydeae.
Getting haploids of wheat, by breeding wheat and corn (Barelay, 1975; Inagaki;, 1985, 1988; Comeau et al., 1988 Ahmand, Cameau, 1990 -- cited by Giura, Cerc Genet Veg Anim III, 17-26 1994) indirectly proved that the genus Zea is able to pollinate with the genus Triticum. So, in corn phylogeny it has been possible for species belonging to other tribes and genera (Andropogoneae - Manisurus; Gramineae - Triticum) to contribute to the appearance and development of the genus Zea, especially to the species Zea mays L., by gene introgression.
Getting a corn mutant through somatic embryogenesis at the Agricultural Research Station, Simnic - Craiova, Dolj, Romania (Urechean, Com Pers Ses Anu Ref ICCPT Fundul, 1996) having characters much more different than the species Zea mays seems to represent the lost transpecies that Mangelsdorf, Reeves and Galinat theoretically placed at the beginning of the species Zea mays as well as Tripsacum dactyloides.
This report shows a series of theoretical considerations, starting with the expression of some phenotypic characters obvious in the resulting mutant as scientific arguments for the possibility of recovering some latent genetical information within the genome of Zea mays by somatic embryogenesis.
Caryopses belonging to the genotype S3 x A188 were collected 10 days after pollination and were sterilized in 70% ethyl alcohol and 1% mercuric chloride (HgCl2). The excised immature embryos inoculated on MS callus medium (Murashige and Skoog, 1962), 30 g/l saccharose, 2 mg/l 2,4-D; pH 5.8, were incubated in darkness at 27-28 C. Passage 1 was done on NBM medium (N6 macroelements, Chu, 1978; B5 microelements, Gamborg, 1968; M vitamins, Murashige and Skoog) without hormones with 12 g/l saccharose, pH 5.8, callus cultivation at a photoperiod of 16 h light and 8 h dark, at 28-30 C. Rooting was done on MS medium with 2.0 mg/l 2,4-D and 30 g/l saccharose, followed by transfer on sterile soil and cultivation in vegetation plots under laboratory conditions. One single seedling of 50 transferred tillers on root medium survived to maturity, representing the mutant described in this work.
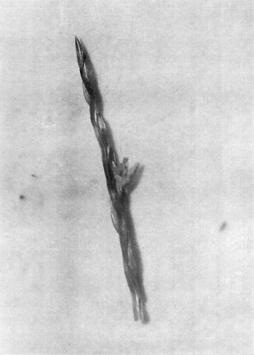
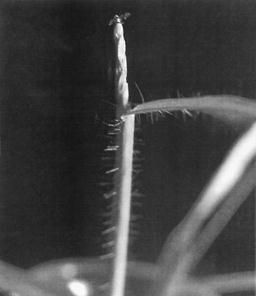
Using the chromosomal maps of Neuffer, Jones and Zuber (Mutants of Maize, Crop Sci Soc Amer, 1968) and those filed by Micu (Genet Issled Kuk, Stiint Chis 211-225, 1981), the known mutants whose phenotypic expression was obvious in the case of the mutant, "hermaphroditic grassy small plant", were identified. The mutant is phenotypically characterized as follows: miniature grassy plant, 8-10 cm height, showing tillers and branches like teosinte type (Figure 1); stem with short internodes, lacking the characteristic concavity at the female flower insertion area; tassel with simple, hermaphroditic flowers (Figures 2, 4); seeds like caryopses having small dimensions 0.6/0.3 mm, not viable, completely closed in glumes (Figure 3).
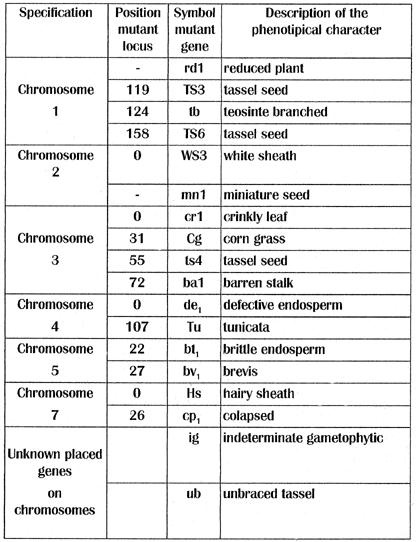
The appearance of a mutant can be induced by: somaclonal variability that appears in somatic embryogenesis, induced by the culture medium; simultaneous modifications, synchronous in one direction, belonging to a large number of genes (Table 1); activation of some transposons within chromosomes 1, 2, 3, 4 and 7 induced by culture stress (Chomet et al., EMBO J 6:295-302, 1987; Schwartz;, PNAS 86, 1989) or epigenetic heredity mechanisms that modify the expression of a large number of homeotic genes (involved in morphogenesis), by modification of methylation patterns.
The mutant phenotypically recalls the genera Manisuris, Andropogonea as well as Coix, Tripsacum and even Triticum by its flowers, spikelets and seeds, differing basically from the current species, Zea mays. The mutant represents a far ancestor of Zea mays L. kept latent in the species genome and reactivated through somatic embryogenesis. The appearance of this mutant opens again the scientific hypothesis concerning the origin and phylogenetic dynamics of corn and might provide scientific proof that there was an hermaphroditic ancestor in the origin of Zea mays, confirming the hypothesis of Mangelsdorf, Reeves and Galinat. The ancestor would be the starting point for the phylogenetic tree from which the current crop corn has evolved. The species Zea mays L. seems to have been developed based on certain sudden macromutations; the morphological-physiological changes incurred became hereditary and entropically preserved.
Table 1. Genetical expression modifications of Zea mays L. in the mutant "hermaphroditic grassy miniature plant".
Figure 1. Hermaphroditic grassy miniature plant.
Figure 2. Terminal hermaphroditic flowers.
Figure 3. The mutant seeds -- caryopses enclosed in glumes and nude seeds compared to a normal wheat seed (x 5).
Figure 4. Leaves with hairy sheath; uniaxial tassel; hermaphroditic simple flowers.
Return to the MNL 71 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}