Heterochromatic knob-specific repeated sequence is associated with
the formation of chromosome bridges in cultured cells and in germinating
roots of aged seeds
-- Fluminhan Jr., A; Ohmido, N; Fukui, K; Kameya, T
The behavior of chromosomes in anaphase cells of embryogenic calli (Type II cultures) has been analysed by means of fluorescence in situ hybridization (FISH) with the 180-bp highly repeated DNA sequence found to be a major component of maize heterochromatic knobs (Peacock et al., PNAS 78:4490-4494, 1981; Dennis and Peacock, J. Mol. Evol. 20:341-350, 1984). Configurations showing the delayed segregation of sister-chromatids, considered to be an initial event in the development of bridges (MNL 66:87-88, 1992; Fluminhan and Aguiar-Perecin, in press), were hybridized in situ with the probe pZm4-14 (kindly supplied by Dr. James Birchler, Univ. of Missouri). Plasmid DNA with the insert was used as the template for direct-labelling during PCR amplification. Biotin-labelled probes were hybridized at 37 C for 8 hours, in a 2xSSC / 50% formamide solution, after heating at 70 C for 6 min., according to Fukui et al. (Theor. Appl. Genet. 87:893-899, 1994). After staining with fluorescein-isothiocyanate (FITC)-avidin conjugate, signals were amplified by applying a biotinylated anti-avidin solution, followed by incubation with a fluorescein-avidin solution. Chromosomes were then counter-stained with a DAPI solution, and examined by fluorescence microscopy. Images were captured by a cooled CCD camera (Photometrics) mounted on the microscope. Digitized images were photographed by a color image recorder.
FISH with selected anaphase configurations confirmed the involvement of the knob-specific repetitive sequence with the event of delayed segregation of sister-chromatids (Figure 1). This observation seems to correspond to the hypothesis described by Phillips et al. (Proc. 7th Intl. Cong. Plant Tissue Cell Cult., pp. 131-141, 1990), that variation in DNA methylation could be a principal factor in the occurrence of chromosome breakage in tissue cultures. Dennis and Peacock (1984) have reported that the 180-bp repeats could show up to ten CG or C_G regions among the different clones sequenced. Since these sites are recognized to be particularly susceptible to methylation of the cytosine, the detection of methylated bases by in situ procedures, as reported in mammalian cells (Miller et al., Nature 251:636-637, 1974), represents an interesting aspect for future investigation.
Furthermore, we have analysed chromosomal aberrations arising during the first mitosis in root tips germinated from long-term stored seeds of different genotypes. Configurations showing the initial event of delayed segregation of sister chromatids have been analysed by FISH as described above. The results were very similar to those observed in cultured cells (Figure 2). This observation suggests that both systems (culture in vitro and storage of dried seeds) could be under the influence of common or related mechanisms of cellular senescence, which would lead to the occurrence of apparently identical cytological abnormalities at mitosis, as discussed elsewhere (Fluminhan and Kameya, Theor. Appl. Genet., in press).
The behavior of broken chromosomes through successive cell divisions has also been investigated. We have collected evidence indicating the occurrence of breakage-fusion-bridge cycles in both systems. Figure 3 shows a late anaphase observed in our studies with cultured cells, with one single bridge containing two hybridization sites to the knob-specific sequence. This figure could have originated from successive B-F-B cycles, as illustrated in Figure 4. Analysis of such configurations by FISH with the telomeric repetitive sequences (TTTAGGG)n has supported the proposed mechanisms. A complete report on these findings is in preparation.
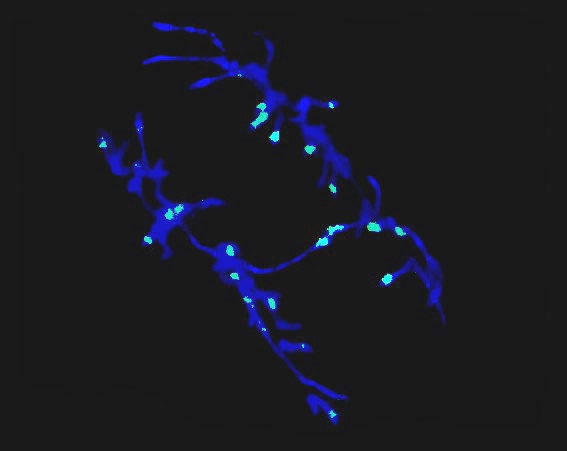
Figure 1. Anaphase cell of an embryogenic callus of S5 progeny obtained from cv. Mexico Amber Kernel after fluorescence in situ hybridization (FISH). The arrows show hybridization sites to the knob-specific repeated sequence on the initial event of delayed segregation of sister chromatids.
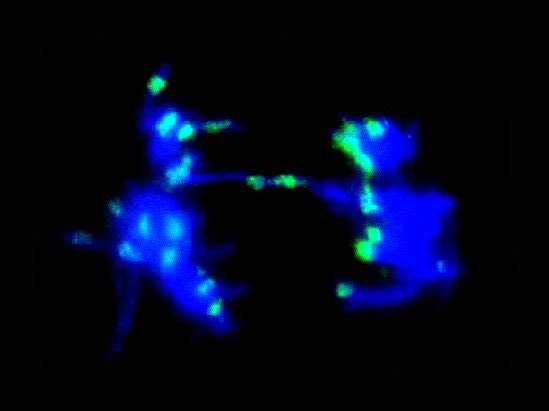
Figure 2. Anaphase cell of germinating roots of aged seeds after FISH. The arrows show hybridization sites on a configuration of delayed segregation of sister chromatids.
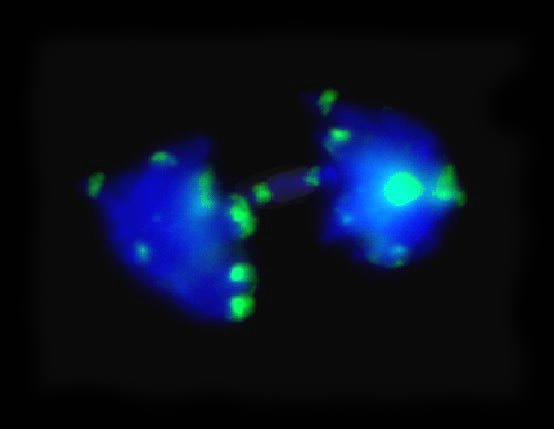
Figure 3. Late anaphase of cultured cells after FISH with the knob-specific repeated sequence. The arrows show hybridization sites on a chromosome bridge possibly originated from successive breakage-fusion-bridge cycles.
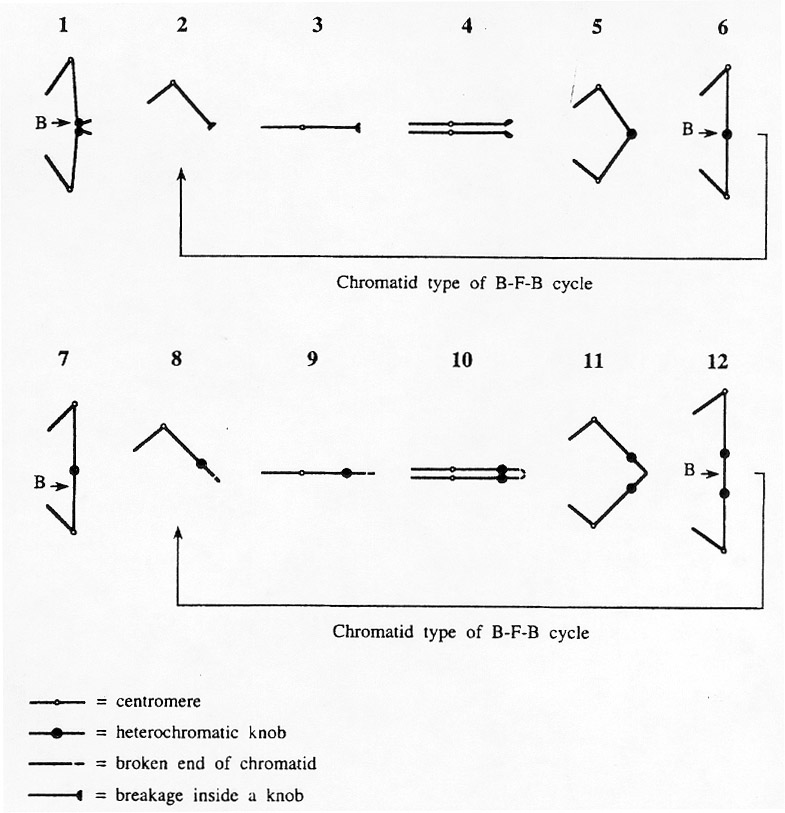
Figure 4. Diagram illustrating the origin of a broken chromosome at mitotic anaphases in cultured cells and in root tips from germinating aged seeds of maize, and its subsequent behavior. 1. Bridge configuration resulting from the initial event of delayed segregation of sister-chromatids (Figures 1 and 2). 2. Primary breakages frequently occur inside the knob. 3. One of the sister cells receives a deficient chromosome with a freshly broken end. 4. Fusion of replicated broken ends during the subsequent mitosis. 5 and 6. A dicentric chromosome is formed that will undergo the chromatid type of breakage-fusion-bridge cycle during each successive nuclear division, conforming to the behavior described by McClintock (PNAS 25:405-416, 1939; Genetics 26:234-282, 1941) with the analysis of gametophyte tissues. 7. Bridge configuration resulting from a previous round of breakage-fusion-bridge. 8. Breakage between the centromere and the knob of the dicentric. 9. The chromosome containing the knob and a freshly broken end is sent to one of the sister cells. 10. Fusion of the replicated broken ends. 11. A dicentric with two knobs is formed (Figure 3). 12. Breakage at different locations gives rise to diverse deficient-duplicated chromosome types. Letter B indicates positions of breakages. A further discussion about these mechanisms is presented in Fluminhan and Aguiar-Perecin (in press) and in Fluminhan and Kameya (Theor. Appl. Genet., in press).
Please Note: Articles submitted to the Maize NewsLetter cannot be cited without the permission of the authors.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}