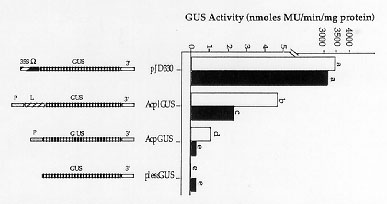
Mobility of the maize Ac-Ds transposable element family depends on the production of Ac-encoded transposase (TPase), a DNA-binding protein which recognizes internal sites near both Ac termini. TPase binding sites at the 5' subterminal region were mapped at or near sequences which may be important for transcription activation (Kunze and Starlinger, EMBO J. 8:3177, 1989). The proximity between the TPase binding sites and the transcription start site led us to hypothesize that TPase may regulate its own transcription, as was found for other transposable elements. This hypothesis was tested in tobacco, in transgenic plants and in protoplasts transformed with different fusions of Ac promoter and leader sequences to a ß-glucuronidase (GUS) reporter gene. The activity of the Ac promoter, from nucleotide 1 (at the 5' termini of Ac) to 346, and Ac promoter and leader (1-960) was determined using plasmids pAcpGUS and pAcplGUS respectively (see Fig. 1). Plasmid pJD330 (35S-GUS) was used both as a positive control for GUS expression and as a control for the TPase effect on the expression of a non-Ac-related promoter. In addition a promoter-less GUS gene (plessGUS) was used as a negative control (Fig. 1). A TPase-encoding construct (St-Ac) was made by subcloning the TPase gene under the regulation of a 35S promoter. All plasmids were built both in a bluescript backbone for use in transient assays and in binary vectors in order to obtain tobacco transgenic plants.
DNA from each of the GUS-fusion plasmids, or calf thymus DNA, was transformed
into tobacco protoplasts, with and without the TPase producing plasmid
pSt-Ac. GUS activity was determined in protoplast extracts, by the fluorimetric
assay with MUG substrate. The average GUS activity of six replicates per
treatment is shown in Figure 1. In treatments without the TPase-producing
plasmid we found that the Ac promoter-leader is ~800 fold weaker
than the 35Sý promoter, as deduced from GUS activity obtained with
pAcplGUS and pJD330 respectively. When GUS-carrying plasmids were co-transformed
with the TPase-producing plasmid, pSt-Ac, we observed a significant reduction
in GUS activity, of two fold with pAcplGUS, and four fold with pAcpGUS
(Fig. 1). On the other hand, the activity of the 35S promoter in pJD330
was not affected by the TPase (Fig. 1). Therefore, we show that TPase can
repress specifically Ac promoter expression, independently of position
effects.
In order to determine the effect of TPase on Ac promoter activity
in transgenic plants, crosses were done between T1 plants carrying the
TPase producing clone (pSt-Ac) and T1 transformants carrying the GUS gene
in which transcription was driven by Ac promoter and leader (pAcplGUS).
The genotype of the progenies from three independent hemizygote T1 pAcplGUS
plants X T1 pSt-Tpase was determined by Southern blots and GUS activity
was measured for each of the 15-20 F1 sibling plants grown in each cross.
In such an analysis, sibling plants which have identical genetic origin
and background and identical genetic dosage of the pAcplGUS construct,
differ only with regards to the presence of the TPase gene. Kanamycin resistant
F1 siblings segregate for the following genotypes in equal ratios: (pAcplGUS/_
, _/_) : (pAcplGUS/_, TPase/_) : (_/_, TPase/_). The effect of TPase on
the element promoter was expressed as the percentage of GUS activity of
double hemizygote plants carrying both GUS and TPase constructs compared
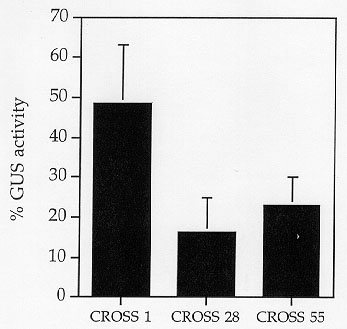
to siblings carrying only the GUS-fusion gene. In the three crosses, we
found a reduction in GUS activity in the presence of the TPase gene, from
two fold in cross 1 to 6.5 fold in cross 28 as shown in Figure 2. Plants
expressing only the TPase ( _/_ , TPase/_ ) had GUS activity similar to
background levels (data not shown). Similar results were found independently
by J. Jones (personal communication). Taken together our and Jones's results
suggest that the TPase-mediated repression observed occurs at the transcriptional
level rather than post transcriptionally.
Figure 1. Transposase effect on GUS expression in transient assays. GUS activity is shown following transformation of tobacco protoplasts with constructs pJD330, pAcplGUS, AcpGUS or plessGUS, without (white columns) or with (black columns) transposase produced from clone pSt-Ac. Treatments with different letters (a to e) are different (P<0.05) as determined by a multiple ranking test (SAS, 1990).
Figure 2. Transposase effect on Ac promoter--sibling comparisons. The transposase effect on GUS expression was studied in young leaves of F1 plants, in three crosses between pAcplGUS and TPase-producing pSt-Ac parents (cross 1, 28, 55). GUS activity of double hemizygote plants carrying both GUS and TPase constructs (as determined by Southern blots) is expressed as a percentage of the GUS activity in siblings hemizygous for the pAcplGUS construct only. The average GUS activity of the pAcplGUS siblings (the 100% value), was 9, 488, and 248 nanomole MU/mg protein/min, for cross 1, 28 and 55 respectively. Standard errors of the means are represented by bars on top of each column.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}