Evolution of Ac/Ds transposable elements
--Rubin, E and Levy, AA
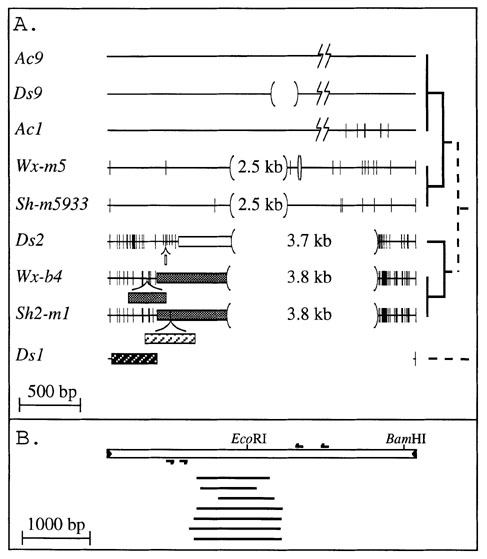
Ac/Ds transposable elements constitute a family which comprises several members (Banks et al. C. S. H. Symp. Quant. Biol. 50:307-311, 1985). Only two Ac elements have been characterized so far, Ac1 and Ac9, which are almost identical in sequence. They both have imperfect terminal inverted repeats (TIRs) and differ in the range of sequencing errors. Ds elements, on the other hand, form an heterogeneous group, both in structure and sequence. A transposable element has been traditionally defined as a Ds based on genetic properties rather than on molecular data: any element which cannot mobilize itself, but can be mobilized by Ac is considered as a Ds element. Subsequent characterization of Ds elements at the molecular level done in several labs is summarized in Figure 1A.
Three kinds of Ds elements can be identified: 1- those with virtually no homology to Ac except in the TIRs, like the Ds1 element (also known as rUq) ; 2- elements with internal deletions, including Ds9 and the Ds in Wx-m5 ; 3- Ds's containing both deletions and insertions in the internal part of the element, including Ds2, and the Ds's from Wx-b4 and Sh2-m1. The double Ds element from Sh-m5933 is made of two identical Ds elements, very similar to that found in Wx-m5, inserted one within the other (Doring et al., Nature 307: 127-130, 1984). All Ds elements, except Ds1, share extensive similarity with Ac, indicating a common origin. However, the mechanism by which they were derived from Ac is not known.
In order to better understand the underlying mechanism of Ds element formation, we have screened for de novo formation of Ds elements in transgenic tobacco, which offers a Ds-free environment. We have used PCR with the primers shown in Fig. 1B and genomic DNA template from plants transformed with Ac or with a Ds element which differs from Ac only by a 4bp insertion (constructs pAGS4411 and pAGS4081 given by H. Dooner). Internal deletions (Fig. 1B) were obtained only with Ac-containing template but not with Ds or with a stable Ac. This suggests that internal deletion formation is transposition dependent and probably occurs by abortion of an Ac-induced gap repair. No insertions have been identified, but we are currently using different primer sets to test for such de novo events.
While only two almost identical Ac elements are known, an increasing number of sequences related to Ac-encoded transposase, from maize and from other distant species, is being reported in sequence databases. These sequences usually come from other known distantly related transposons. Other sequences, with unknown functions, have been reported as Ac-related. No "host" genes with a known function have been found so far, with homology to Ac. This suggests that Ac comes from a superfamily of ancient transposons rather than it being a recent host gene which became mobile. The family of Ac-related elements had been originally designated as the hAT family, for hobo, Tam3 and Ac (Calvi et al., Cell 66: 465-471, 1991), other elements were added to this family as summarized by Essers and Kunze (MNL 69: 39-40, 1995). Here we report on further extension of Ac-related sequences with the addition of two transposons, Hermit and Hector (Fig. 2), and four sequences of unknown function. Two of these sequences, from C. elegans, share only one block of homology with all the other sequences. This block (see sequence in "conserved region III" by Essers and Kunze MNL 69:39-40, 1995) might be "the hAT-box" common to all members of the hAT superfamily. Two other sequences, namely the maize and rice ESTs, have been recognized as entries in the plant sequences database and were further sequenced in our lab. The definition of conserved blocks in the hAT superfamily might be helpful to better understand transposase functions and to further extend the superfamily to other species.
Figure 1. (A) Classification of known Ds elements. Ds elements for which sequence data are available (with the exception of Ds9, for which only a high-resolution restriction map is available) were compared to Ac. Deletions, compared to Ac, are shown in parentheses. Small polymorphisms are indicated as vertical lines. Blocks of insertions are shown as boxes. Boxes with the same filling are related. The phylogenetic tree, describing relatedness but not distances among Ds elements, is shown on the right, rooted on the unknown progenitor of the Ac/Ds family. It is based on a minimal evolution tree derived from the sequences of the subterminal regions at the left and right ends of the element, which are shared by all elements. In Sh-m5933, two elements, identical to the one shown here are inserted one within the other. (B) Nested PCR was used to detect de novo Ds elements formed in transgenic tobacco plants. Primer position is indicated by small arrows above (forward primers) and below (reverse primers) a schematic representation of Ac. The position and size of each deletion relative to Ac was determined by sequencing, and is shown as a bold line. Primer length is not drawn to scale.
Figure 2. The hAT superfamily. Sequence databases were screened for similarity to Ac, Hobo or Tam3 using iterative DBase searching. Alignments were performed using the Macaw package, using both the GIBBS and diagonal parsing algorithms. Sequences whose gene symbol is in italics are thought to be transpositionally active. For some sequences, obtained through genome projects of different species, no function was assigned (Mest, Rest, celc10a4 and cek09al1). For Rest alignment was done with preliminary sequencing data obtained in our lab. Boxes with similar filling are homologous. Horizontal lines indicate unique sequences.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}