The clusters of development genes as seen against the UMC 1995 map
--Khavkin, E and Coe, EH
Previously (MNL 68:61, 1994; MNL 69:106, 1995; Russ. J. Plant Physiol. 42: 408 & 558, 1995) we have hypothesized that corn developmental genes associate into functionally meaningful clusters, 10 to 30 cM long, comprising the loci for environmental and hormonal sensors (e.g., phy1, abp1, d8 and vp1), the growth machinery genes (e.g., for the enzymes of hormone synthesis) and the master genes presiding over the spatial and temporal transitions in cell growth and development (e.g., homeobox genes). The initially delineated clusters accounted for most of the naked eye polymorphisms related to growth and development, including les and nec loci presumably associated with programmed cell death. The clusters that manifested the most comprehensive pattern of developmental genes usually included knox and/or other homeobox sequences. The majority of over 400 major QTLs for plant architecture, growth and development in vivo and in vitro, the grain yield as the integer of growth, and ABA accumulation and effects, mapped within these clusters.
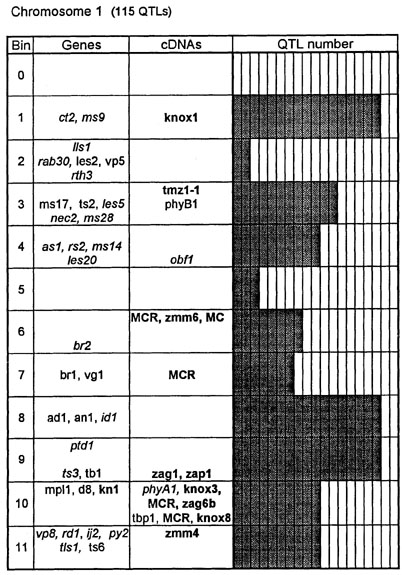
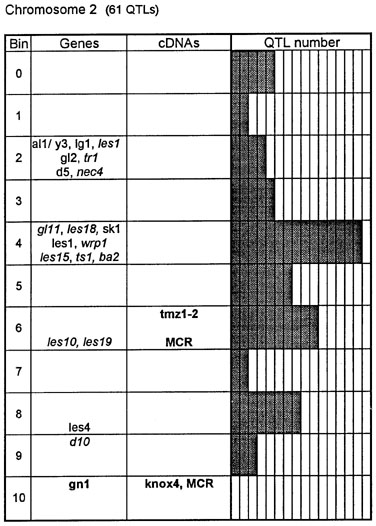
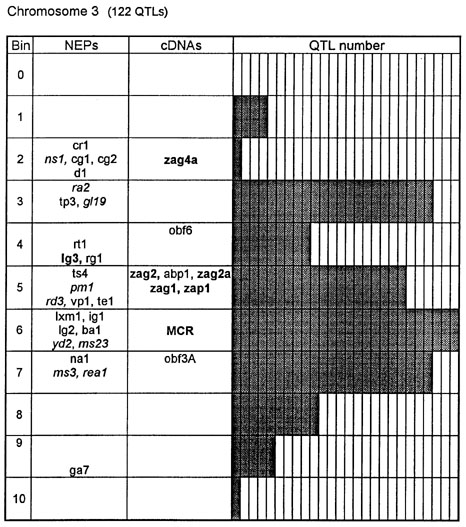
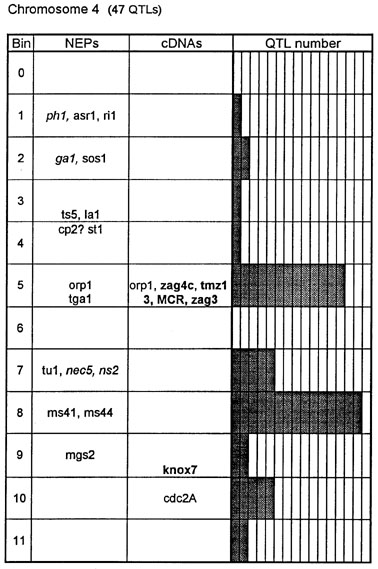
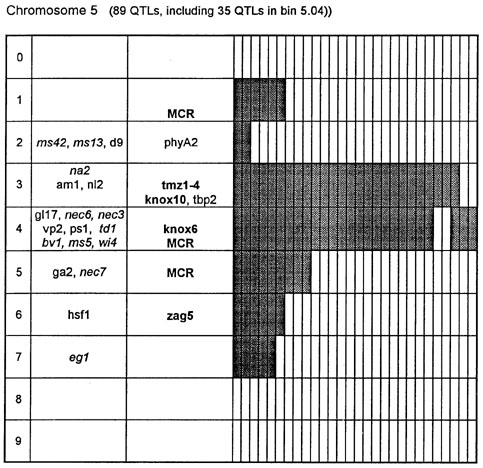
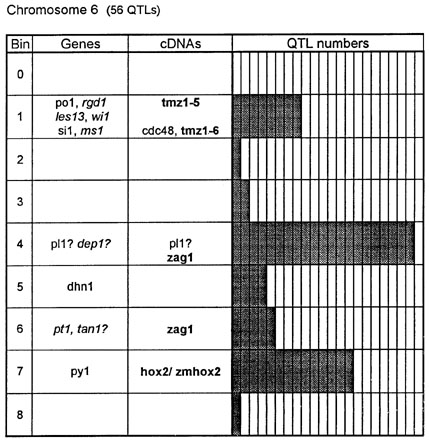
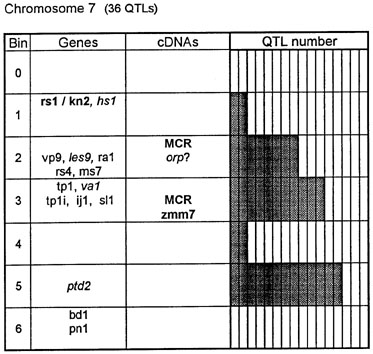
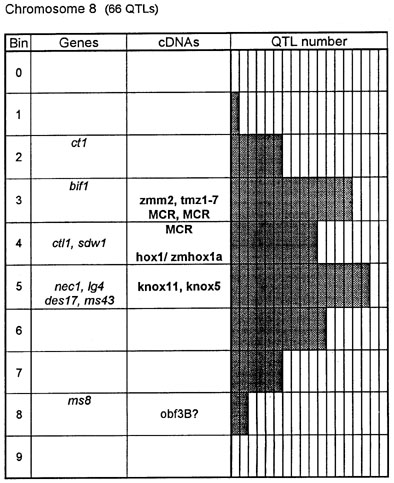
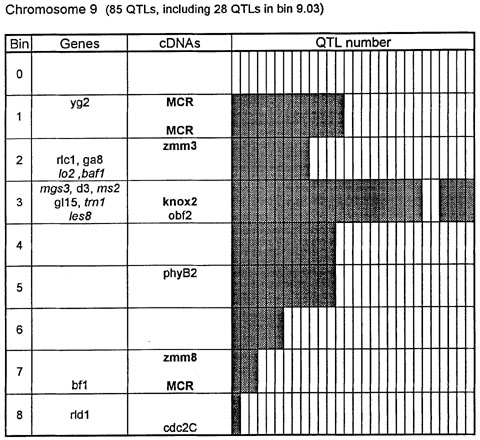
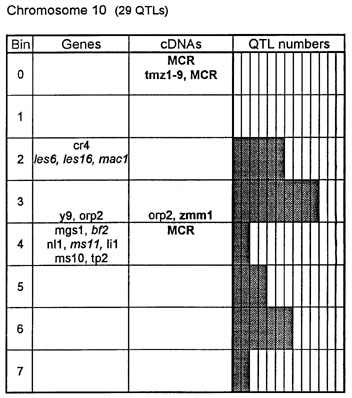
Figure 1. Chromosome 1 (115 QTLs), chromosome 2 (61 QTLs), chromosome 3 (122 QTLs), chromosome 4 (47 QTLs), chromosome 5 (89 QTLs, including 35 QTLs in bin 5.04), chromosome 6 (56 QTLs), chromosome 7 (36 QTLs), chromosome 8 (66 QTLs), chromosome 9 (85 QTLs, including 28 QTLs in bin 9.03), chromosome 10 (29 QTLs). Genes and QTLs for growth and development. Tentatively located genes and sequences are italicized; homeobox genes and sequences are in bold. MCR are MADS-box containing RFLPs. Coinciding MADS-box sequences mapped in different laboratories could represent one and the same locus.
Figure 1 presents the profiles of the developmental genes, cDNA sequences and QTLs refitted to the new UMC 1995 map. Several new genes, e.g. for gametophyte development, and new homeobox sequences were added using the data from the 1995 Gene List, 1995 UMC and BNL maps and the papers by Kerstetter et al. (Plant Cell 6:1877, 1994), Fischer et al. (Nucl. Acids Res. 23:1901, 1995) and Mena et al. (Plant J. 8: 845, 1995). The QTL database was supplemented from several additional sources (CIMMYT, 1994 QTL data in MaizeDB; Abler et al., Crop Sci. 31:267, 1991; Schön; et al., Heredity 70:648, 1993; Doebley et al., J. Hered. 85:191, 1994; Ragot et al., Crop Sci. 35:1306, 1995; Ajmone-Marsan et al., Theor. Appl. Genet. 90:415, 1995; Austin and Lee, MNL 69:7, 1995; Beaumont et al., Genome 38:968, 1995; Berke and Rocheford, Crop Sci., 35:1542, 1995).
Taking into consideration the mapping accuracy, the clusters of developmental genes generally coincide with the location of homeobox sequences and with the QTLs for growth, development and grain yield, especially in chromosomes 1, 3, 5, 8 and 9. The most prominent and challenging exceptions are the bins 2.02 and 4.07, where only a few QTLs correspond to several important developmental genes, and homeoboxes are missing, while in the bins 6.04 and 7.05/06, the largest QTL peaks and/or several homeobox genes are inadequately matched by naked eye polymorphisms; in contrast, in the bins 2.10 and 10.00 homeobox genes are not substantiated with the classical developmental genes and QTLs.
When accepted as a working model, the cluster hypothesis poses several questions which at present can be answered only tentatively, by referring the reader to other relevant evidence. (1) What are the putative functional and physiological advantages of gene association into clusters? We presume that the clusters are the units of genes expressed in concert to contribute for plant growth, development and apparently some of the plant responses to stress. The close association of the functionally related genes in the clusters would contribute to compartmentation of signal molecules and help cooperatively recruit the transcription factors, e.g. MADS-box proteins, into multicomponent regulatory modules of high specificity (Krumlauf and Gould, Trends Genet. 8:297, 1992; Jacob, C. R. Acad. Sci. 316:331, 1993; Shore and Sharrocks, Eur. J. Biochem. 229:1, 1995) and thus would facilitate fine tuning of growth and development. (2) Why are several physiologically different traits of plant development mapped by one and the same molecular probe? We may envision QTLs as projections onto the phenotype of the key structural loci providing for the various essential elements of growth and development (dwarf and viviparous genes are good examples) or of the master switches of development, like knox and MADS-box genes, and thus such loci are pleiotropic by definition. (3) Why is one and the same developmental trait mapped to several widely distant loci? Two answers are possible. First, the loci defined as different genes can manifest one and the same physiological trait (e.g., stunted growth). Second, drawing an analogy from metabolic regulation, we may believe that the position of a bottleneck locus in one and the same developmental pathway may change in different genotype x environment interactions, and thus different key genes are manifested in various segregating populations employed for QTL mapping. (4) Why so many clusters? One partial answer to the evident redundancy of developmental clusters is the hypothesis of paleopolyploid corn origin; in addition, later duplication events could contribute to the redundancy: it is remarkable that most clusters border the centromeres where duplicated regions are most often found (Helentjaris, MNL 69:67, 1995). (5) What is the adaptive significance of developmental gene associations? A suggestion to support the advantage of clusters comes from the evidence of selective pressure maintaining the polygenic complexes that comprise relatively few pleiotropic genes (e.g., for plant height and flowering control) as the integral units through the evolution of Poaceae (Lin et al., Genetics 141:391, 1995; Paterson et al., Science 269:1714, 1995); the superiority of clusters may reside in the complementary gene interaction within a conserved chromosome segment resulting in numerous manifestations of heterosis (Bingham et al., Crop Sci. 34:823, 1994). (6) How do different genes for physiologically similar functions (e.g., plant height) and the whole clusters as functional units interact when redundant and located on different chromosomes? The recent topic of homology-dependent gene silencing may hopefully provide some explanation in the near future.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}