The dappled mutants affect endosperm development
--Castiglioni**, PF; Allegra**, D; Hoxha*, M; Todesco**, G; Dolfini**,
S and Gavazzi*, G
While screening for mutants affecting aleurone pigmentation we isolated several independent mutants exhibiting a mutable aleurone pattern. The mutants are referred to as Dap (defective aleurone pigmentation) and their origin, segregation values and kernel weight are given in Table 1. Two additional isolates, Dap*-9 and Dap*-10, have not yet been analyzed.
Table 1. Dappled mutants description.
|
|
|
|
|
|
|
|
|
|
||||
| Dap*-1 | EMS | 43.1 | 1.0 |
|
|
| Dap*-2 | EMS | 40.9 | 2.3 |
|
|
| Dap*-3 | EMS | 43.9 | 1.2 |
|
|
| Dap*-4 | UV | 36.3 | 6.1 |
|
|
| Dap*-5 | XRAYS | 38.7 | 1.9 |
|
|
| Dap*-6 | EMS | 47.8 | 2.1 |
|
|
The phenotype of Dap seeds consists of purple tissue of variable size and shape on a yellow background, as previously described by Stinard and Robertson (MNL 61:7-9, 1987). In addition, opaque white sectors are frequently observed on the yellow background. Dap*-6 can be easily distinguished from the other isolates, since it conditions dark purple sectors on a weak red background as if pelargonidin rather than cyanidin is accumulating. All six Dap mutants are associated with a significant reduction in seed size, leading sometimes to extremely defective seeds.
The segregation values reported in Table 1 are observed when Dap/+ females are crossed to purple aleurone males. In fact, a common feature shared by all Dap isolates is the observation that crosses in which Dap plants are used as females segregate for colored and dappled seeds, while crosses in which Dap plants are used as males give only colored seeds. These segregations disclose a significant shortage of the mutant over the expected one-half and a dominant expression of the mutant over the wild-type allele.
Although dappled is not expressed in male outcrosses, it is male transmissibile in the case of Dap*-3, Dap*-4 and Dap*-6, but not of Dap*-1, Dap*-2 and Dap*-5 isolates. Its recovery, however, is erratic and lower than the expected one-half (compare results obtained in 1994 and 1995 in Table 2). So, when Dap/+ plants enter the cross as males, in three cases Dap gametes contributing to the endosperm formation are apparently selected against; on the contrary, in the other three isolates male transmission leads to gene silencing in the endosperm, since the mutant is recovered and expressed again in the next generation, if transmitted through the female.
Table 2. Male transmission of Dap as determined in the progeny
ears from outcrosses of heterozygous dappled males to purple aleurone females.
Results obtained in summer 1994 and 1995 are presented separately.
| Mutant symbol |
|
|||
| 1994 | n(1) | 1995 | n(1) | |
| Dap*-1 | none | 53 | none | 61 |
| Dap*-2 | none | 31 | ND(2) | |
| Dap*-3 | 44.4 | 54 | 18.2 | 66 |
| Dap*-4 | 34.2 | 38 | 14.7 | 34 |
| Dap*-5 | none | 49 | none | 70 |
| Dap*-6 | 16.6 | 78 | none | 58 |
The reduced frequency of Dap seeds recovered in both male and female outcrosses and the lack of recovery of homozygous Dap seeds in the selfed progeny of Dap/+ heterozygous plants could be the result of a gametophytic selection against Dap alleles operating more drastically in the male versus female gametophytic generation. If selection is mediated by chromosome breaks, pollen sterility should be observed and indeed the field analysis of plants heterozygous for Dap*-1, Dap*-2, Dap*-3 and Dap*-6 confirms the expectation (estimated pollen sterility of 30% or more).
To inquire about Dap mutants and their transmission, it would be useful to establish their chromosomal location. We first attempted to obtain this information with Dap*-1 and Dap*-2, because we had some indication of their linkage with sh1, a marker of the short arm of chromosome 9. The recombination values of the three point testcross of heterozygous Dap Bz Sh / + bz sh females to homozygous + bz sh males are 17.7 ± 1.7 for Dap-bz and 2.9 ± .7 for bz-sh (n = 490). To establish if Dap is proximal or distal to the sh-bz segment a three point testcross was done with bz and wx (bz-wx distance 25 cM). The results place Dap proximal to bz, leaving still undefined the orientation of Dap in regard to wx. Assuming Dap is distal to wx the recombination values of the bz wx + / Bz Wx Dap testcross are 18.0 ± 1.6 for bz-wx and 2.1 ± .6 for wx-Dap respectively (n= 606). Dap*-2 also appears to be located on the short arm of chromosome 9 (Dap*-2-sh recombination value : 18.3 ± 1.2; n = 1000), suggesting a possible allelism between the two mutants.
To test the allelism of Dap mutants, one can cross different Dap isolates inter se, select dappled seeds in the F1 and outcross F1 females to purple aleurone stock. One-third of the ears so obtained should be homozygous Dap in the case of allelism or segregate Dap vs. colored seed in a 3:1 ratio in the case of non allelism and independent assortment; if the two Dap are linked, one-third of the progeny ears should segregate a majority of Dap seeds (75% or more, depending on the linkage intensity).
Out of seven outcrosses of Dap*-1/Dap*-3 F1 females, five segregate Dap and colored seeds with a Dap shortage (37%), while two ears show an excess of Dap seeds (66.9%), a result expected in case of non-allelism. Progeny ears of Dap*-1/Dap*-4 female outcrosses segregate Dap and colored seeds with a shortage of Dap. This unexpected result could indicate allelism, if we assume that Dap*-1/Dap*-4 seeds are not viable. For a cytological characterization of the mutant, different approaches were followed. Histological sections were obtained from seeds of different Dap mutants, at 25 days after pollination and after an exposure to light for 48 hours. The presence of a continuous aleurone layer was observed in all mutants, demonstrating that the lack of anthocyanin accumulation in colorless sectors is not the consequence of the absence of aleurone cells in the depigmented areas. The same conclusion was reached by scanning electron microscopic analysis on the Dap*-1 mutant.
In order to correlate cellular morphology with presence or absence of pigments, fixation and histological procedures were performed preserving anthocyanins in the aleurone cells. Colorless aleurone cells show an abnormal morphology, if compared to pigmented cells. In general, colorless cells appear irregular in morphology, smaller and flatter than normal ones and occasionally binucleate. More than one layer of aleurone cells may be present and sub-aleuronic cells are irregular and disconnected. The defect is confined to cells of the endosperm, since histological sections of mature seed embryos do not reveal any difference between normal and mutant seeds.
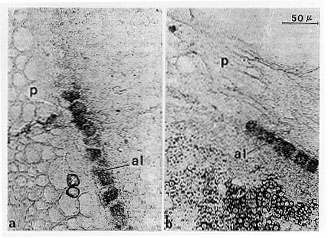
To investigate the lack of pigment in aleurone cells, in situ hybridization experiments were set up, with the aim to check for the presence of A1 mRNA in colored and colorless regions of the aleurone. A1 is a structural gene coding for dihydroquercetin reductase, an enzyme of the pathway which leads to anthocyanin accumulation. The experiments show a correlation in all Dap mutants between the presence of A1 mRNA and the presence of anthocyanins in colored aleurone cells. On the other hand, in cells devoid of pigments, the A1 transcript seems absent (Fig. 1). These results would demonstrate that the anthocyanin biosynthetic pathway in mutant cells is blocked and could help in further studies on this mutation.
Figure 1. a and b. Presence of A1 transcripts exclusively in colored aleurone cells of Dap*-2 seeds (25 DAP) visualized by in situ hybridization; al aleurone, p pericarp.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}