Studies on the genetic control of apomixis in Tripsacum
--Grimanelli, D; Leblanc, O; Perotti, E; González-de-León,
D and Savidan, Y
Apomixis in higher plants refers to several mechanisms of asexual reproduction through seeds. In all cases, apomictic processes bypass both meiosis and egg cell fertilization, producing offspring which are exact genetic replicas of the mother plant. In Tripsacum (x=18), the closest apomictic relative of corn, all polyploids reproduce through the diplosporous type of apomixis (Leblanc et al., Am. J. Bot. 82:57, 1995). Diplospory results from meiotic failure in megasporocytes that directly develop into mature unreduced female gametophytes through three or more mitoses. Typically, it is a facultative phenomenon, and an apomictic plant usually produces both asexual (apomeiotic) and sexual (meiotic) embryos. Apomixis in Tripsacum, as in other apomictic species studied, has been thought to be controlled by one dominant allele.
As part of an effort to transfer apomixis to maize from Tripsacum, we recently reported the identification of RFLP markers linked with diplospory in a maize-Tripsacum F1 population (Leblanc et al., TAG 90:1198, 1995). We used the markers to analyze various generations of maize-Tripsacum hybrids and backcross derivatives and define a model for the genetic control and inheritance of diplosporous reproduction. Here we report some results and propose a transposon-tagging-based strategy for further studies of apomixis in Tripsacum.
As expected, maize and Tripsacum genomes are significantly colinear. This is obviously of great interest for mapping Tripsacum: we can switch from anonymous mapping to comparative mapping, and screening for a specific zone in the Tripsacum genome can be based efficiently on maize mapping information.
An important feature of apomixis is its relation to polyploidy: except for rare exceptions, apomicts are polyploids, while sexuality in the same species, if known, is usually found at lower ploidy levels. Two hypotheses have been proposed regarding the mechanism thought to prevent apomixis in diploid genotypes. The first one (Nogler, Bot. Helv. 92:13, 1982) assumes that the allele or alleles responsible for the apomictic development are not transmitted through haploid gametes. Therefore, apomixis would not be recovered in diploid plants. The second one proposes a dosage effect in which diploid plants do not express apomixis, although the corresponding alleles may be transmitted (Mogie, Biol. J. Lin. Soc. 35:127, 1988; Noirot, J. Evol. Biol. 6:95, 1993). Our results (Grimanelli et al., manuscript in preparation, summarized in Fig. 1) suggest that the gene(s) controlling apomixis in Tripsacum are linked with a segregation-distorter-type allele promoting the elimination of the "apomixis allele(s)" when transmitted through haploid gametes. This would explain why apomixis is not recovered in diploid plants, and has strong implications for transferring apomixis to diploid crops.
Furthermore, a significant difference appeared when we looked at the segregation of four RFLP loci surrounding the gene of interest in both the apomictic tetraploid and the sexual diploid Tripsacum. It seems that recombination is restricted at the tetraploid (apomictic) level as opposed to both the diploid (sexual) in Tripsacum and maize, as represented in their RFLP maps.
Because the specific chromosome segment shows a restricted level of recombination, the classical model of monogenic inheritance for apomixis probably needs more careful analysis: whatever the number of genes involved, they surely behave as a single locus in segregating populations. This observation is consistent with the existence of a segregation distorter linked with the "apomixis allele(s)". Meiotic drive systems can usually be associated with chromosomal structural modifications, such as inversions, that locally restrict recombination, further creating linkage disequilibrium between the distorter allele, the target loci, and eventually modifier alleles (for a review of segregation distorters, Lyttle, Ann. Rev. Gen. 25:511, 1991).
Several on-going programs are aiming at the isolation of the genetic system responsible for apomixis, and its transfer to crops. The usual hypothesis is that a single allele could account for the whole developmental process of apomictic reproduction. Our results do not claim the existence of several genes, but at least suggest the possibility of a cluster of linked loci. To determine the number of genes controlling apomixis, as well as potentially to isolate the corresponding alleles, we started a transposon tagging experiment. Apomictic maize-Tripsacum dihaploids (10 chromosomes of maize + 18 chromosomes of Tripsacum) were crossed to Mutator lines, kindly provided by Mike Freeling. Because apomixis is essentially a facultative phenomenon, most of the progenies are clones of the mother plants, but about 10% result from fertilization of unreduced gametes. Those plants consequently have 20 chromosomes of maize plus 18 of Tripsacum, and are both apomictic and Mu active. We are presently checking the level of Mu transposition.
This transposon tagging experiment has three major objectives. First, we expect to obtain evidence for the existence of regulatory activities: while the plants remain perfectly apomictic, the level of expression of the trait (level of facultativeness) may vary, due to disruptions of some regulatory genes. Second, from a qualitative point of view, we may disrupt apomictic development totally or partially, and therefore get information concerning the number of genes involved. A major target in that case is the segregation distorter allele, since it represents a strong limitation for the transfer of apomixis to maize. Finally, we should be able to analyse the behavior of Mu when transmitted through ameiotic gametes.
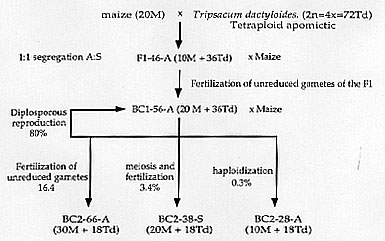
Figure
1. Chromosome numbers, constitution and modes of reproduction in maize-Tripsacum
hybrids and some backcross derivatives (A: apomictic, S: sexual, M: maize,
Td: Tripsacum dactyloides x=18). Under the hypothesis of Mendelian
inheritance of apomixis, the character is expected to segregate 1:1 in
the BC2-38 plants. Of ca. 6000 progenies we produced from the facultative
apomictic BC1 plants, 218 BC2-38 plants originated from the sexual development
of embryos, but none was apomictic. By contrast, rare dihaploid plants
were produced through parthenogenetic development of such reduced gametes,
and were found to be apomicts. Segregation of the diagnostic bands for
mode of reproduction is consistent with the expression of the trait. The
segregation indicates a strong selection against the apomictic allele(s).
The analysis of further generations (data not shown) suggests that the
distortion is best explained by the presence of a segregation distorter
allele, linked with apomixis.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}