Lost locus resurfaces? The possible involvement of brown pericarp1
in determining silk maysin concentration
--Byrne, PF; McMullen, MD; Snook, ME; Musket, T; Widstrom, NW; Wiseman,
BR and Coe, EH
Concentration of maysin, a C-glycosyl flavone, in maize silks is an important resistance factor against the corn earworm, Helicoverpa zea (Boddie). Because maysin synthesis occurs as a branch of the flavonoid metabolic pathway, our research has sought to identify and estimate the contributions of loci from that pathway that affect maysin levels.
In a study of the population (GT114 x GT119)F2, silk maysin concentrations of 285 plants were measured with reversed-phase HPLC. RFLP analysis was conducted on the same plants, using probes encoding flavonoid pathway enzymes or linked marker loci, a total of 39 loci distributed on all chromosomes except chromosome 8. Single-factor analysis of variance was used to detect significant associations between maysin concentration and genotypic classes at individual RFLP loci, based on a comparison-wise error rate of 0.05. Epistasis was evaluated by testing the significance of all possible pairwise combinations of loci (excluding closely linked loci) in two-way analyses of variance.
Last year (MNL 69:53-54) we reported the results of our analysis to date: major effects on maysin concentration were associated with the p1 region of chromosome 1 (accounting for 58% of the total phenotypic variance) and the c1 - bz1 region of chromosome 9 (accounting for 6.7% of the variance). We were uncertain which locus in the latter region affected maysin levels, but felt that c1 was a likely candidate because of its partial homology with p1, its similar role as a pathway regulator, and testcross results indicating different c1 alleles in GT114 and GT119.
To better estimate the position of the responsible locus on chromosome 9, we probed for additional loci on either side of the c1 - bz1 region, namely umc109, umc105a, wx1, and csu147. Analysis of variance showed a peak in percent variance explained (10.8%) at umc105a, midway between bz1 and wx1. This position is close to the reported location of brown pericarp1 (bp1) (Meyers, Ohio J. Sci. 5:295-300, 1927; Emerson et al., Cornell Univ. Agric. Exp. Stn. Memoir 180, 1935). The homozygous recessive condition at that locus together with a functional p1 factor for pericarp color was reported to result in the production of brown pigmentation in the pericarp in place of red phlobaphenes. Though bp1 was identified and mapped over 60 years ago and included on linkage maps for many years, stocks containing the mutation have apparently been lost, and the locus was removed from working maps in 1983. We believe that the locus detected in our study by umc105a may be bp1 for the following reasons:
(1) The agreement in chromosome location, approximately midway in the interval between bz1 and wx1.
(2) Identical interactions with the p1 locus. In our study, umc105a affected maysin concentration only when it was homozygous recessive and a functional p1 allele was present, i.e., only when the pathway was activated by p1. These are the same conditions required for observation of the brown pericarp phenotype.
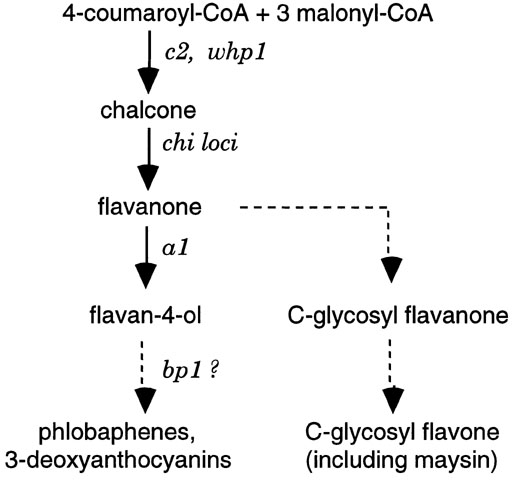
(3) Parallels with a1-controlled brown pericarp. Recessive a1 plus dominant p1 produce brown pericarp and is reported to also enhance accumulation of C-glycosyl flavones, the class which includes maysin, in pericarp and silks (Styles and Ceska, Can. J. Genet. Cytol. 19:289-302, 1977; Styles & Ceska, Maydica 34:227-237, 1989). A block at the a1 step in the pathway leading to phlobaphenes and 3-deoxyanthocyanins (Fig. 1) presumably leads to a build-up of flavanones and other intermediates, some of which are then shunted into the flavone branch of the pathway. Because of the similarities of effects, the site of action of bp1, like that of a1, seems likely to be in the pathway leading from flavanone to 3-deoxyanthocyanins and phlobaphenes.
To determine whether GT119, the source of the umc105a allele conditioning higher maysin values, carries a recessive allele at bp1, we plan to cross the line to a red pericarp stock. F2 progeny that segregate for brown pericarp would support, but not prove, a recessive bp1 allele in GT119. To conduct additional experiments we continue searching for an existing bp1 stock; we would appreciate hearing from MNL Cooperators having such a stock or additional information about bp1.
Figure 1. Part of the p1-controlled portion of the flavonoid pathway in maize. Loci are shown in italics. Dashed lines indicate steps that are less well characterized or that require more than one enzyme.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}