TnpA is a multifunctional DNA binding protein encoded by the maize Suppressor-mutator (Spm) transposable element. TnpA is required for transposition and is both a repressor of the unmethylated Spm promoter and an activator of the methylated promoter. While analyzing the protein using a yeast GAL4-based hybrid system in transiently transformed tobacco cells, we found that TnpA represses the >10-fold transcriptional activation observed when the GAL4 DNA binding domain is used alone. By contrast, a 33- to 45-fold activation of the Spm promoter was observed when the VP16 activation domain was tethered to TnpA. TnpA binding sites, but no TATA box, were required for transcription activation. Among the TnpA deletion derivatives tested, those retaining the coding sequences for the DNA-binding and protein-dimerization domains gave the highest level of transcription activation when fused with the VP16 activation domain. As shown below, the TnpA gene and TnpA binding sites in the short Spm promoter therefore provide a novel, highly sensitive single-hybrid system for identifying and studying plant transcription activation domains in plant cells.
The full-length TnpA coding sequence and 5'- and 3'-terminal deletion
derivatives of TnpA were fused to the yeast GAL4 DNA-binding domain and
tested for their ability to affect transcription of a LUC reporter gene
expressed from a minimal plant promoter containing GAL4 binding sites (Figure
1; Giniger et al., Cell 40: 767-774, 1985; Trentmann et al., Mol. Gen.
Genet. 238: 201-208, 1993; Schläppi et al., Cell 77: 427-437, 1994).
The Herpes Simplex VP16 activation domain fused to the GAL4 DNA-binding
domain served as a positive control (Triezenberg et al., Genes Dev. 2:
718-729, 1988). The effect of the fusion genes was compared with the baseline
transcriptional activation observed with the GAL4 binding domain in plant
cells. Effector plasmid DNAs carrying the various translational fusions
were coated onto tungsten particles together with a GAL4 binding site-containing
LUC reporter plasmid (Fig. 1B) and introduced into tobacco suspension cells
by microprojectile bombardment (Russell et al., In Vitro Cell Dev. Biol.
28: 97-105, 1992; Raina et al., Proc. Natl. Acad. Sci. 90: 6355-6359, 1993).
A plasmid containing a bacterial CAT gene expressed from the CaMV 35S promoter
was used to normalize for transformation efficiency. LUC and CAT activities
were assayed 18 hours after introduction of the DNA and the ratio of LUC
to CAT activity was calculated to correct for between-experiment variation.
The LUC/CAT ratios obtained with different constructs were then compared
with the relevant control by calculating the ratio of the LUC/CAT value
obtained with an experimental construct to that obtained with the relevant
control construct and expressing the ratio as "relative activation." Thus,
for example, in Figure 2A, the reference control value is the LUC/CAT ratio
obtained with an antisense effector plasmid, while in Figure 2B, it is
the LUC/CAT value obtained with an effector plasmid expressing the GAL4
DNA-binding domain.
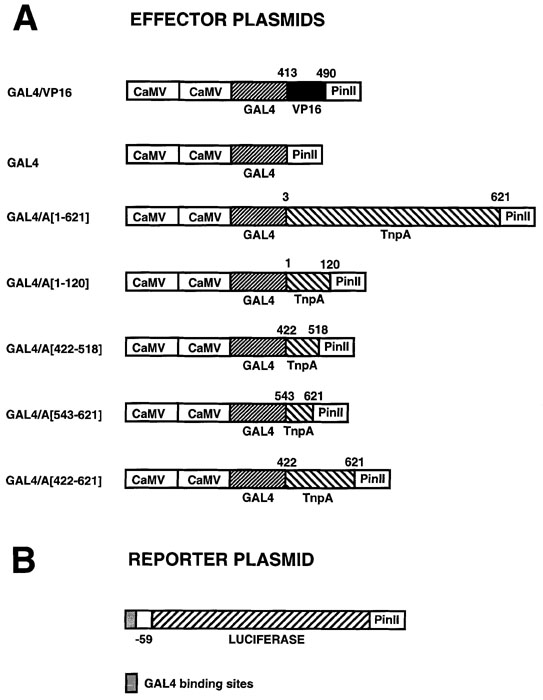
Figure
1. Schematic representation of GAL4 fusion genes and the luciferase
reporter gene.
(A) Effector plasmids are those that encode a trans-acting activator or repressor of the luciferase reporter gene. Each construct contains the yeast GAL4 DNA binding domain either alone or translationally fused to the following: VP16, the strong activation domain of the Herpes Simplex virus VP16 (amino acids [a.a.] 413 to 490); A[3-621], full-length TnpA (a.a. 3 to 621); A[1-120], N-terminus of TnpA (a.a. 1-120); A[422-518], the protein dimerization domain of TnpA (a.a. 422 to 518); A[543-621], C-terminus of TnpA (a.a. 543 to 621); A[422-621], the dimerization domain and C-terminus of TnpA (a.a. 422 to 621). PinII, potato proteinase inhibitor II terminator; CaMV, Cauliflower Mosaic Virus 35S promoter.
(B) The reporter plasmid contains 5 GAL4 DNA binding sites upstream from a truncated CaMV promoter (bp -59 to +2) and the firefly luciferase gene.
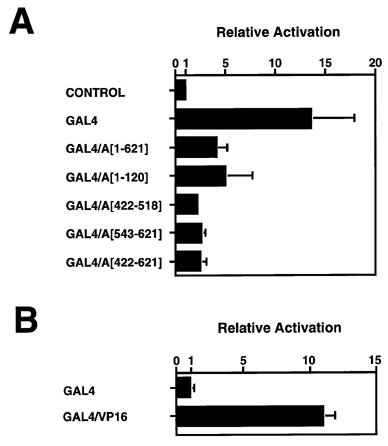
Figure
2. Expression of a GAL4-LUC reporter gene in the presence of GAL4-TnpA
and GAL4-VP16 effector plasmids.
(A) Relative activation of the LUC reporter gene in tobacco suspension cells by expression of the chimeric GAL4/TnpA genes shown in Figure 1. The activity of the test plasmid was measured and normalized to an internal CAT control as described in Experimental Procedures. Relative Activation represents the ratio of normalized LUC activity obtained in the presence of a GAL4- or a GAL4/TnpA effector plasmid to that observed with an effector plasmid carrying an antisnese GAL4/VP16 construct. The LUC reporter plasmid contains GAL4 DNA binding sites and a minimal CaMV 35 S promoter (Figure 1B).
(B) Activation of the LUC reporter gene by the chimeric GAL4/VP16 construct shown in Figure 1. Relative Activation is the ratio of normalized LUC activity observed in the presence of the GAL4/VP16 fusion to that observed with GAL4 binding domain alone.
A plasmid containing the coding sequence for the GAL4 DNA-binding domain activates the LUC reporter gene 14-fold (Fig. 2A; relative activation: 14.3 ± 4.4) in tobacco cells. All of the chimeric genes containing TnpA coding sequences fused to a sequence encoding the GAL4 DNA-binding domain showed lower levels of LUC expression than that detected with the GAL4 DNA-binding domain sequence alone (Fig. 2A). While these observations are consistent with our previous report that TnpA represses its own promoter (Schläppi et al., Cell 77: 427-437, 1994), the fact that even small segments of the coding sequence are inhibitory suggests that their addition to the GAL4 binding domain simply serves to interfere with its ability to interact with other proteins. When the strong VP16 activation domain is fused to the GAL4 DNA-binding domain, expression of the reporter gene is stimulated only an additional 10-fold over the background value observed with the GAL4 DNA-binding domain alone (Fig. 2B; relative activation: 10.9 ± 0.9). Thus the GAL4-based system is relatively insensitive in plant cells because of the high basal activation observed with the GAL4 DNA-binding domain alone.
As previously reported, TnpA represses its own promoter (Cook and Fedoroff, MNL 66: 11-12, 1992; Schläppi et al., Cell 77: 427-437, 1994). The Spm promoter is short (0.2 kb) and contains 9 12-bp TnpA binding sites (Gierl et al., EMBO J. 7: 4045-4053, 1988; Raina et al., Proc. Natl. Acad. Sci. 90: 6355-6359, 1993). To determine whether TnpA can be converted from a repressor into an activator by addition of a strong activation domain, the coding sequence of the VP16 activation domain was fused to different deletion derivatives of the TnpA coding sequence (Figure 3A). TnpA and TnpA/VP16 fusion gene plasmids were co-bombarded into tobacco suspension cells with reporter plasmids in which the LUC gene was expressed from the Spm promoter (Fig. 3B). In contrast to the GAL4 DNA binding domain, which itself stimulates LUC expression substantially (Fig. 2), TnpA constructs lacking VP16 have no detectable background activity . Instead, expression of TnpA constructs containing the DNA-binding and dimerization domains represses the weak Spm promoter, as previously reported (Figure 4A, insert; Schläppi et al., Cell 77: 427-437, 1994).
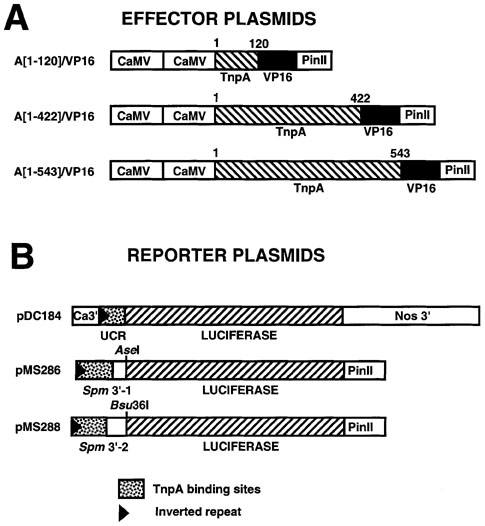
Figure 3. Schematic representation of chimeric TnpA-VP16 and Spm promoter-luciferase reporter genes.
(A) Effector plasmids contain translational fusions of the following TnpA domains to the Herpes Simplex VP16 activation domain (a.a. 413 to 490): A[1-120], N-terminus of TnpA (a.a. 1 to 120); A[1-422], N-terminus and DNA-binding domain of TnpA (a.a. 1 to 422); A[1-543], N-terminus, DNA-binding and protein dimerization domains of TnpA (a.a. 1 to 543). PinII, potato proteinase inhibitor II terminator; CaMV, Cauliflower Mosaic Virus 35S promoter.
(B) The firefly LUC gene in the reporter plasmids is expressed from either the UCR promoter sequence of the Spm element, which contains 9 TnpA binding sites, (Raina et al., Proc. Natl. Acad. Sci. 90: 6355-6359, 1993), or 1 of 2 different fragments of the Spm 3'-end in the antisense orientation , each containing 15 TnpA binding sites (Masson et al., Genetics 177: 117-137, 1987; Gierl et al., EMBO J. 7: 4045-4053, 1988). Nos 3', nopaline synthase terminator. Ca3', CaMV 35S terminator; PinII, potato proteinase inhibitor II terminator.
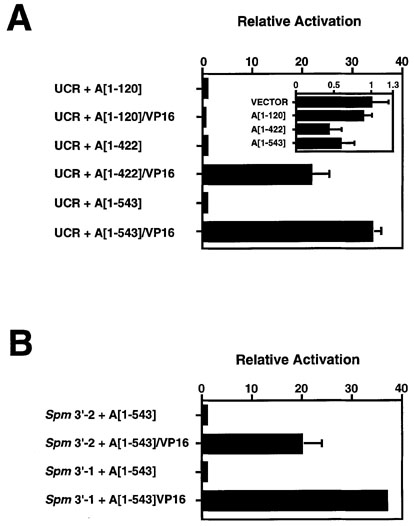
Figure 4. Activation of the Spm Promoter by TnpA/VP16 fusion proteins in tobacco cells.
(A) Relative activation of UCR-LUC reporter gene expression (Fig. 3B) by the chimeric TnpA/VP16 fusion proteins shown in Figure 3A. Controls were plasmids carrying the corresponding TnpA gene or gene fragments lacking the VP16 activation domain. LUC activity was measured and normalized as described in Experimental Procedures. Relative Activation is the ratio of normalized LUC activity observed in the presence of the effector plasmid expressing the TnpA/VP16 to that observed with an effector plasmid expressing only the corresponding TnpA gene or gene fragment. The inset shows the corresponding relative activation values for the TnpA gene fragments relative to the bluescript pKS(+) vector control.
(B) Relative activation of 1 µg Spm 3'-LUC reporter plasmid cobombarded into tobacco suspension cells with 1 µg effector plasmid containing chimeric TnpA/VP16. Fold Activation is expressed as the ratio of TnpA/VP16-effected relative promoter activity to background activity effected by TnpA alone. The Spm 3'-end contains 15 TnpA binding sites. The constructs are shown in Figure 3.
Expression of plasmids carrying certain TnpA-VP16 fusions activates transcription of the LUC gene from the Spm promoter (Figures 3 and 4). A fusion of the VP16 activation domain to the first 120 amino acids of TnpA does not activate expression of the LUC gene (Figure 4A), while VP16 fusions containing the TnpA DNA binding domain do. A VP16-TnpA fusion protein which contains the TnpA binding domain, but lacks the protein dimerization domain, stimulates promoter activity 21-fold over that observed with TnpA alone (Figure 4A, A[1-422]/VP16). The VP16 fusion gene containing both the DNA-binding and the protein dimerization domains of TnpA is the strongest activator (Figure 4A, A[1-543]/VP16). Relative to A[1-543] alone, the A[1-543]/VP16 fusion activates the Spm promoter more than 30-fold (relative activation: 34.0 ± 1.79). The range of promoter activation varied between 33- and 45-fold in different experiments. Thus the addition of an activation domain to TnpA converts it to a strong activator of the Spm promoter.
Two observations suggest that the TnpA binding sites are the most important determinant of the Spm promoter's response to the VP16-TnpA fusions. First, the element's 3'-end, whose sequence organization resembles that of the 5'-end and contains 15 TnpA binding sites (Fig. 3B; Masson et al., Genetics 177: 117-137, 1987; Gierl et al., EMBO J. 7: 4045-4053, 1988) can substitute for the Spm promoter in the present assay. As shown in Figure 4B, the A[1-543]/VP16 fusion activates expression of the LUC gene from two different Spm 3'-end fragments by 20- to 40-fold. Second, the Spm promoter is a TATA-less promoter (Raina et al., Proc. Natl. Acad. Sci. 90: 6355-6359, 1993) and addition of a TATA box does not further enhance the ability of a VP16-TnpA fusion protein to activate the LUC gene from the Spm promoter (data not shown). Because TnpA is normally a repressor of the unmethylated promoter, the baseline or background activity observed with the Spm promoter-driven reporter gene is extremely low, providing a highly sensitive plant-specific system for detecting and analyzing transcription activation domains of proteins. In the present experiments, LUC activity was 33-45 times the background value using the VP16/TnpA/Spm promoter hybrid system, as compared with only about 10-fold over background with the VP16/GAL4 system in plant cells. While the absolute value of the activation was lower for the Spm promoter-based hybrid protein system, the higher sensitivity of the system permits detection and analysis of much weaker activation domains than the GAL4-based system. In addition, this plant-based hybrid system may permit detection of transcription activation domains which require plant-specific co-factors.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}