Spm element: Significance of multiple TnpA binding sites
--Raina, R and Fedoroff, N
The maize Suppressor-mutator (Spm) transposable element encodes two proteins, TnpA and TnpD, which are necessary and sufficient for transposition (Masson et al., Plant Cell 3:37, 1991). TnpA affects the epigenetic state of the Spm element by activating the methylated inactive Spm promoter (Schlappi et al., Cell 77:427, 1994). TnpA is a DNA-binding protein (Gierl et al., EMBO J. 7:4045, 1988) and there are multiple copies of its 12-bp binding site located at the element's 5' and 3' ends. However, the role of TnpA and TnpD in the transposition of Spm is not understood. We have previously reported that TnpA binds to the ends of the element in a concentration-dependent manner (Raina and Fedoroff, MNL 69:13, 1995). We hypothesize that once TnpA is bound to its binding sites, higher order protein-protein interactions bring the ends of the element together. Here we analyze the effect of the presence of multiple TnpA binding sites on binding of TnpA and interaction of DNA-TnpA complexes.
To assess whether the presence of multiple binding sites at the ends of the Spm element is important for the formation of higher order complexes, we studied the binding of TnpA to various deletion derivatives of the 5'-end of the element. The derivatives have 1, 2, 3, 6 and 9 binding sites. Over-expression and purification of TnpA in E. coli have been described previously (Raina and Fedoroff, MNL 69:13, 1995). Target DNAs with 1 to 9 binding sites (except for one with 2 binding sites) were generated by exonuclease III deletions of the 5'-end of the element (Raina et al., Proc. Natl. Acad. Sci. USA 90:6355, 1993). DNA fragments with 2 binding sites were generated by cloning an oligonucleotide corresponding to TnpA binding sites 2 and 3 at the 5'-end of the element in the EcoRV site of bluescript KS+. The fragment was released by digestion with enzymes BamHI and PvuI.
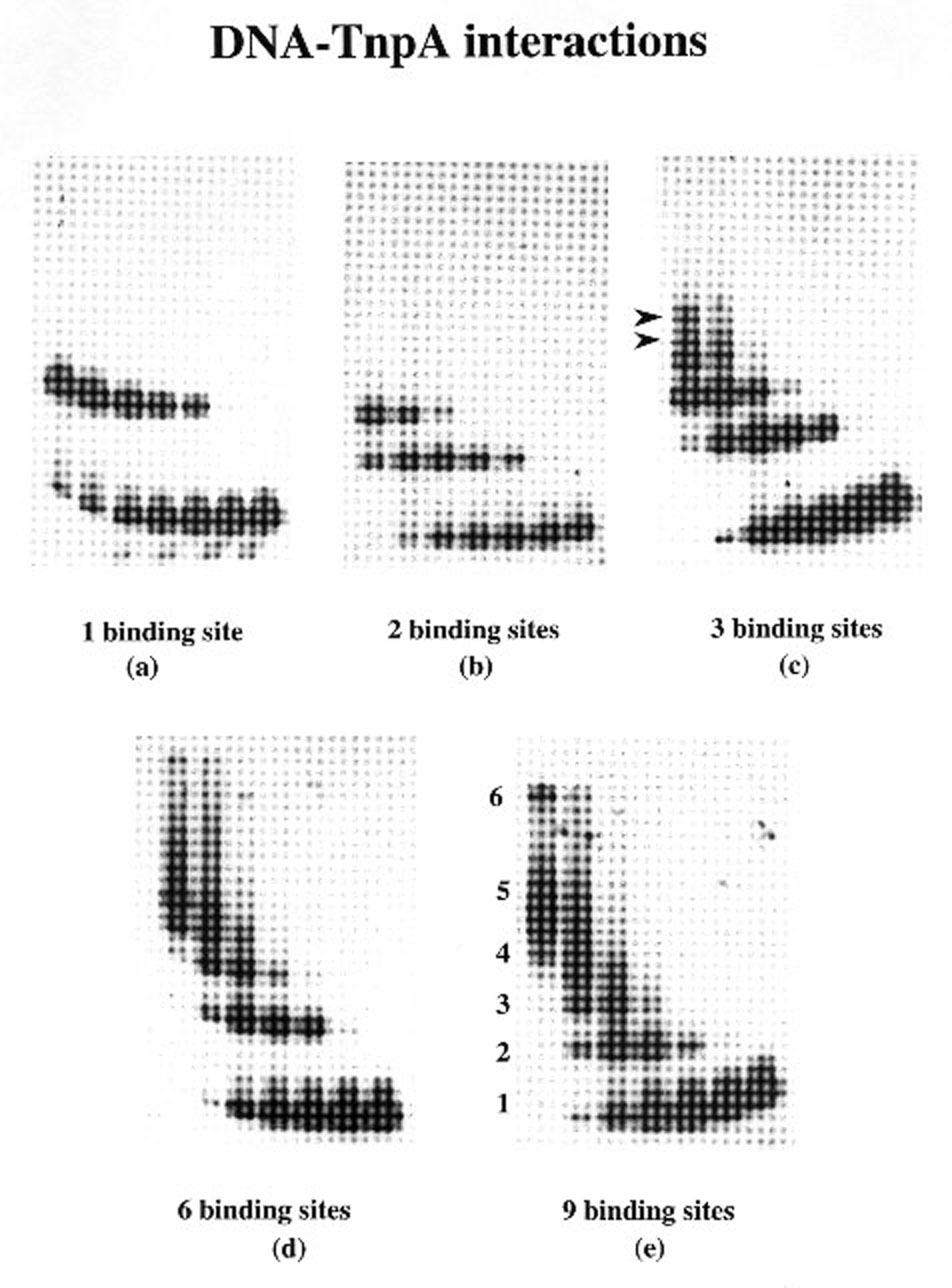
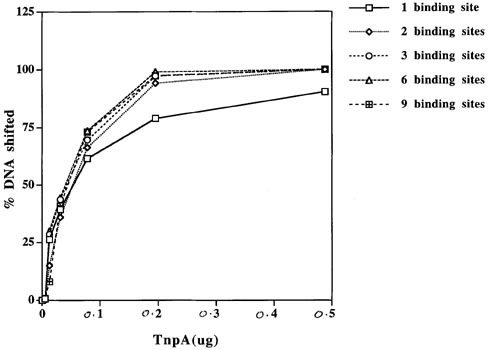
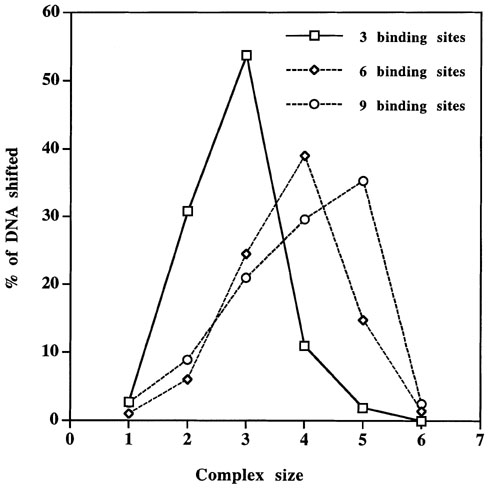
The effect of multiple binding sites on binding of TnpA to DNA was studied by a band-mobility shift assay. The results are shown in Figure 1. The same amount of labeled DNA and protein has been used in all the experiments. The DNA was labeled at both the ends in all cases except for the fragment with 2 binding sites, in which only one end was labeled. The results of this experiment show that the binding of TnpA increases similarly with the protein concentration for all fragments tested. As the protein:DNA ratio increases, more and more sites are occupied (Figures 1 and 2), giving bands of larger size. We have not observed stronger binding of TnpA to a tail-to-tail dimeric binding site than a monomeric binding site, as reported previously (Figure 1a, b; Trentmann et al., Mol. Gen. Genet. 238: 210, 1993). Fragments containing 1 and 2 binding sites give one and two slower-migrating complexes, respectively (Figure 1a, and b). These complexes correspond to one or two sites occupied by TnpA. However when a DNA fragment with 3 binding sites is used in these experiments, we observe more than the expected 3 bands (Figure 1c). When DNA with more binding sites is used in these experiments, we find that a higher fraction of the shifted DNA is in large complexes and this fraction increases with increasing numbers of binding sites per DNA molecule (Figure 3). The large complexes probably arise by intermolecular protein-protein interaction between the TnpA molecules already bound to DNA.
Because we see no evidence of dimerization of TnpA with a fragment containing a single binding site, we propose that the dimerization domain of TnpA is involved in the formation of intermolecular TnpA-DNA complexes. This hypothesis is supported by the observation that fraction of shifted DNA in higher order complexes increases with increasing numbers of TnpA binding sites/molecule (Figure 3). We therefore propose that the dimerization domain of TnpA is involved in protein-protein interactions between TnpA molecules already bound to DNA and that it functions to bring the ends of the element together during transposition.
Figure 1. Band shift assay using deletion derivatives of 5'-end of the element and TnpA.
Figure 2. Graphical representation of data in Figure 1.
Figure 3. Graphical representation of fraction of shifted DNA in each complex. A representative complex size is shown in Figure 1e.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}