As a result of studies conducted on Mutator-induced Bf1 mutants (Bf1-Mu), three bm4 mutant alleles at the bm4 locus were found. All Bf1-Mu mutants produced in these studies were selected against the Bf1-R allele (mutant phenotype - homozygous seedlings, and anthers of heterozygous or homozygous plants, fluoresce blue under U.V. light). Each of the original Bf1-Mu isolates was of the genotype Bf1-Mu/Bf1-R. These original isolates (214 total) were crossed as females to a standard line (bf1+/bf1+), resulting in progeny plants of the genotypes Bf1-Mu/bf1+ and Bf1-R/bf1+. Plants from the progenies of thirty-eight of the isolates, which had been outcrossed to standard, were crossed reciprocally to Bf1-R Bf1-R stocks, as a first screen to determine if any of these isolates might involve deletions. If a deletion was involved, about half of the plants, when outcrossed as males, might have a reduced frequency of Bf seedlings in their outcross progenies. Whereas, these same plants, outcrossed as females, might have about 50% Bf seedlings or less in their outcross progenies. If less, however, the female outcross progenies should have more Bf seedlings than the male outcross progenies. Twenty-seven of the isolates tested showed this pattern of inheritance and were tested further in an attempt to obtain additional evidence that would support the assumption that a deletion was involved.
The putative bm4-Mu mutants occurred in the tests of three of these isolates, Bf1-Mu-044-4, Bf1-Mu-046-6 and Bf1-Mu-546-5. Twenty kernels were planted from the ears of the plants that had reduced transmission of the Bf1-Mu homolog through the pollen in the reciprocal crosses with homozygous Bf1-R stocks. Two genotypes are expected in the progeny plants from these female outcrosses: Bf1-Mu/Bf1-R and bf1+/Bf1-R. If plants of the former genotype were pollinated by homozygous bm4-R plants, and if the Bf1-Mu event was a deletion of sufficient size to include the distal bm4 locus, bm plants would be expected to segregate in the progeny of this cross. The test involving the Bf1-Mu-044-4 isolate resulted in seven out of nine progenies that segregated for bm plants. The expected genotypes in these segregating progenies are Bf1-Mu/bf1+ bm4-R and Bf1-R bm4+/bf1+ bm4-R. The former genotype would result in bm plants with Bf anthers, if Bf1-Mu-044-4 is a deletion that involves both the bf1 and bm4 loci. The latter genotype would result in green plants. A total of 18 bm plants were found in these seven progenies. There are other possible explanations for the results from the reciprocal crosses and the bm4 test other than a deletion that involves both the bf1 and bm4 loci. There are several combinations of two or more simultaneous events induced by the Mutator system that could account for these observations. For example, the Bf1 mutant phenotype could be caused by the insertion of a Mu element at this locus and the bm4 phenotype could be due to a Mutator-induced deletion of this locus. Another possibility is that the Bf1 phenotype is the result of a deletion of this locus and the bm4 phenotype is the result of an insertion mutation. Perhaps both the Bf1 and bm4 phenotypes were the result of the insertion of two different Mu elements and a Mutator-induced deletion, which occurred in the proximity of these loci, but did not include either of them. Because multiple Mutator-induced alterations in one egg involving a short chromosomal region are highly unlikely, the simple explanation of a deletion involving both loci is the most reasonable, until proven otherwise.
The test of Bf1-Mu-046-6 resulted in one family with one bm plant out of five outcross progenies tested. Although nine outcross progenies of Bf1-Mu-546-5 were grown, none segregated for bm plants.
The bm plants from the tests of the first two isolates were pollinated by standard plants, resulting in progeny plants of the following genotypes: Bf1-Mu/bf1+ bm4+ and bf1+ bm4-R/bf1+ bm4+. Plants with Bf anthers, the former genotype, were reciprocally crossed to homozygous Bf1-R plants to determine if the same results were obtained as when the isolate was first tested in reciprocal crosses with homozygous Bf1-R plants.
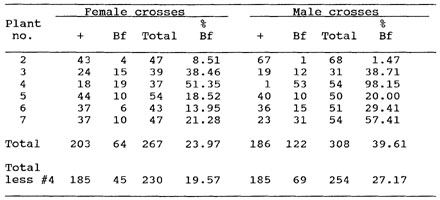
In five out of six reciprocal outcross tests of the Bf1-Mu-044-4 isolate, the results were close to the original reciprocal outcross tests (average values: male outcross progenies 2.04% Bf seedlings and female outcross progenies 37.66% Bf seedlings). The progeny of one of the bm plants of this isolate, however, did not give results that duplicated those of the original reciprocal outcross test. None of the plants gave results (Table 1) that approach those observed in the original or second reciprocal outcross tests of this isolate. Also there was no consistent pattern of inheritance seen for the six plants tested. They vary with respect to the percentage of Bf seedlings in both the male and female outcross progenies. In the progenies of all but one plant (#2) there are more Bf seedlings in the male crosses than the female crosses. This is not a result expected if a deletion were present.
Table 1. Results of seedling tests of reciprocal crosses of the individual plants from the outcross progeny of the first putative bm4-Mu mutant (derived from the Bf1-Mu-044-4 isolate).
These results suggest that the bm plant crossed with standard had a different origin than the rest of the bm plants of this isolate. If the Bf1-Mu/Bf1-R plant pollinated by the homozygous bm4-R plant had an active Mutator system, which is very likely, there is the possibility that it could have produced an egg carrying a Mutator-induced bm4-Mu mutant in the homolog with the Bf1-R allele. When this egg was fertilized by a sperm with the bm4-R allele, a bm plant with the genotype Bf1-R bm4-Mu/bf1+ bm4-R would result. A bm plant that had this origin, when pollinated by standard pollen, would have progeny plants of the following two genotypes: Bf1-R bm4-Mu/bf1+ bm4+ and bf1+ bm4-R/bf1+ bm4+. Plants of the former genotype would have Bf anthers, and when crossed reciprocally to Bf1-R Bf1-R plants would not be expected to give frequencies of Bf seedlings observed in the original tests of the Bf1-Mu-044-4 isolate because they do not carry the putative deletion. Such is the situation observed in Table 1. The erratic transmission, however, observed in this table would not be expected if a simple insertion mutant was responsible for the bm4-Mu allele. (See below for a discussion of this and similar patterns of transmission exhibited by the other two putative bm4-Mu mutants.)
The isolate Bf1-Mu-046-6 probably does not carry a deletion that includes the bm4 locus, because only one bm plant was found in the bm4 test. Further, when plants from the outcross of this isolate with standard were tested in reciprocal crosses with homozygous Bf1-R plants (Table 2) the original transmission pattern of Bf seedlings was not observed (i.e., 46.10% Bf seedlings in the female outcross progeny and 27.04% Bf seedlings in the male outcross progeny). If the Bf1-Mu-046-6 is not a deletion, which includes the bm4 locus, how is the occurrence of this one bm plant explained? It could have occurred in the same manner as the atypical bm plant in the test of the Bf1-Mu-044-4 isolate. The reciprocal crosses of Bf1-R bm4-Mu-046-6/bf1+ bm4+ plants to homozygous Bf1-R plants show an erratic transmission pattern similar to that of the bm4-Mu mutant from the tests of the Bf1-Mu-044-4 isolate.
Table 2. Results of seedling tests of reciprocal crosses of the individual plants from the outcross progeny of the second putative bm4-Mu mutant (derived from the Bf1-Mu-046-6 isolate).
The third putative bm4-Mu mutant resulted from a different crossing procedure than the former two. A plant of the putative genotype Bf1-Mu-546-5/bf1+ [from the cross of the original isolate (Bf1-Mu-546-5/Bf1-R) by a plant from a standard line] was pollinated by a plant heterozygous for the A-B translocation TB-9Lc, which involves most of the long arm of chromosome nine. The same plant, which was pollinated by TB-9Lc, was outcrossed as a male to a homozygous Bf1-R plant. This cross was made to determine if the plant pollinated by the TB-9Lc stock carried the putative deletion, which the results of the original reciprocal cross of this isolate to homozygous Bf1-R plants suggested might be present. In the original test, the male outcross progenies had 27.93% Bf seedlings. In the outcross test of the plant pollinated by TB-9Lc, 30.45% of the seedlings were of the Bf phenotype. On the surface these two percentages appear to be reasonably close. However, the former percentage was statistically different from a 1:1 ratio at the one percent level (n = 111), while the latter was not significantly different from a 1:1 ratio at the five percent level (n = 13). Thus, there was a distinct possibility that the plant pollinated by TB-9Lc was not heterozygous for Bf1-Mu-546-5, but was instead heterozygous for the Bf1-R allele. Because of the small size of the outcross progeny in this test (n = 13) and because the chi square value was close to that expected for significance at the five percent level, the progeny of the cross with TB-9Lc was tested further to determine if this plant was heterozygous for Bf1-Mu-546-5. Two Bf seedlings occurred in the progeny of this cross, which were transplanted to the field. Both of these plants had the phenotype expected for hypoploid TB-9Lc plants (i.e., short plants with narrow leaves, rudimentary tassels that only occasionally extrude anthers that shed no pollen, and small ears, which in most plants produce kernels when pollinated). Unexpectedly, these two plants had the brown midrib phenotype. Because there is no other bm mutant known on the long arm of chromosome 9, these plants probably are hemizygous for a Mutator-induced mutant at the bm4 locus. Only one of these plants produced an ear and it was pollinated by a standard plant. If the original plant pollinated by TB-9Lc was heterozygous for the Bf1-Mu-546-5 allele, all progeny plants of this cross were expected to be of the genotype Bf1-Mu-546-5/bf1+, and should have shown a reduced frequency of Bf seedlings in progenies from the male outcrosses to homozygous Bf1-R plants. The results (Table 3) do not duplicate the Bf1-R test results of the original isolate. Thus, the hypoploid plant probably does not carry the Bf1-Mu-546-5 allele. This would mean that the plant pollinated by TB-9Lc was heterozygous for the Bf1-R allele and the hypoploid Bf seedlings were hemizygous for this allele. The most logical explanation for the origin of the bm phenotype is that it was the result of a Mutator-induced bm4 mutant on the homolog that carried Bf1-R in the original Bf1-Mu-546-5/Bf1-R isolate that was pollinated by standard. The homolog with Bf1-R and the closely linked putative bm4-Mu mutant allele had an erratic transmission pattern (Table 3) similar to that of the previous two bm4-Mu mutants. Note: the latter mutant has not been confirmed as being a mutant at the bm4 locus. It, however, must be on the long arm of chromosome nine and as yet no other mutant with the bm phenotype has been described on this arm. This fact along with its erratic transmission pattern, which is similar to those observed for the other two bm4-Mu mutants, strongly suggests that it is a bm4-Mu mutant.
Table 3. Results of seedling tests of reciprocal crosses of the individual plants from the outcross progeny of the third putative bm4-Mu mutant (derived from the Bf1-Mu-546-5 isolate).
Chi square tests for heterogeneity of the female and male outcross progenies in Tables 1, 2, and 3 were all significant at the one percent level. Why was this erratic transmission pattern observed for all three of these mutants? It would suggest that these Mutator-induced mutants at the bm4 locus must have involved more than simple insertion mutations. Probably they were the result of more complex changes that happened to involve this locus and resulted in the mutant bm phenotype at the same time.
The following are a few unanswered questions about the results reported above: Is it just coincidence that three bm4 mutants derived from different Bf1-Mu isolates happen to have erratic transmission patterns? Could it be that there is something involved in the induction of the Bf1-Mu isolates that is responsible for this unusual behavior of a bm4 mutant, when a mutation occurs at this locus in these stocks? Would bm4 mutants that were induced directly by the Mutator system (i.e., were not derived from a Bf1-Mu stock) show the erratic transmission patterns observed for the mutants in this report? What mechanism is responsible for the transmission pattern of these mutants?
It should be pointed out that the results reported here are based on the transmission, not of the mutant bm4-Mu allele, but on the transmission of the closely linked Bf1-R allele. All plants tested had the putative genotype of Bf1-R bm4-Mu/bf1+ bm4+ and, thus, the transmission of the bm4-Mu allele would be expected to closely approximate that of the Bf1-R allele. There remains the possibility, however, that some kind of recombination event is taking place that eliminates the bm4 mutant allele from the gametes that function and at the same time is responsible for the erratic transmission of the Bf1-R allele. This possibility could be tested by crossing sibling plants from those used for generating the data reported in Tables 1, 2, and 3 with plants homozygous for both Bf1-R bm4-R and simultaneously scoring the progenies for both the Bf and bm phenotypes. Another test could be made by self-pollinating Bf plants from the progenies of the reciprocal crosses of the putative Bf1-R bm4-Mu/bf1+ bm4+ plants. Most of the progenies from these selfs should segregate for bm plants, if no crossing over between the bf1 and bm4 loci had occurred.
I will gladly supply seeds to anyone who is interested in analyzing these mutants further.
Return to the MNL 70 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}