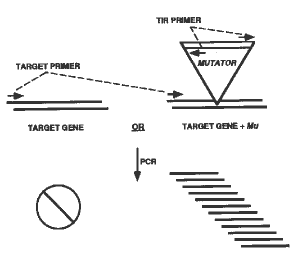
Of the scientific tools available to researchers studying gene function in organisms from other kingdoms, the ability to selectively knock out a gene is paramount. There are many examples where directed mutagenesis can be carried out simply to disrupt or otherwise recover mutations within a specific gene of interest. Accurate interpretation of the mutant phenotype most nearly describes the role of the normal gene. For plant biologists, elucidation of gene function has equal value and priority, but in the absence of needed mutants, it currently requires the use of more cumbersome techniques such as anti-sense or co-suppression, which depend upon the generation of transformants. Transformation is not a limitation for some dicot species, but transgenic methods suffer from a lack of uniformity of suppression and/or a lack of specificity for a single target gene. We have produced a new tool that will alleviate some of the difficulty in studying gene function in plants, specifically, a reverse-genetics system developed for maize. This resource consists of a collection of genomic DNAs from approximately 40,000 F1 plants produced by crossing Mutator-active lines* with inbred/hybrid lines (in effect, a library of the transposon-mutagenized genome), coupled with a collection of the F2 seed from each plant. Mu insertion alleles for a known DNA sequence can be selected from the DNA collection using PCR: a primer specific for the gene of interest is paired with a primer specific for the terminal-inverted-repeat (TIR) region of Mu elements. Amplification of the target gene can occur only if an allele exists in which a Mu element has inserted in or near the gene of interest (see illustration). Prospective insertion alleles are identified by successive rounds of PCR/DNA dot blot hybridization (using a probe specific for the target gene); first on DNA pools, subsequently on DNA from individuals. Once individuals are identified, kernels from the corresponding F2 seed packet can be allocated for phenotypic analysis of specific mutant alleles. To illustrate the utility of such a system, brief examples of its application are useful:
1) To knock out genes. Though many DNA sequences for genes are known (be it from random sequencing, cloning by homology, reverse genetics from peptide sequence, or transposon tagging), mutations in specific genes can be difficult to isolate (e.g. lethal when homozygous, phenotype unknown, masked by a redundant function). From this collection, which has proven to be very rich in mutations, Mu insertion alleles can be selected for any gene whose sequence is known. Though not all insertion alleles will be mutagenic, due to the position or orientation of the Mu insertion, precise knowledge of the gene's structure can be used to selectively target exons, or a particular region of the sequence, in order to maximize the potential for recovering mutagenic insertions. Phenotypic analysis and the propagation of insertion alleles proceeds from the F2 seed, which segregates the mutant and wild-type alleles. Lethal mutations can be discerned by the inability to recover homozygotes, yet heterozygotes can be used to propagate the allele. Gene families or redundant functions can be targeted independently, and insertion alleles combined genetically to observe the net phenotypic effect of stacked mutations. Tissue-specific or conditionally expressed genes, once cloned, can be selected for disruption, facilitating the mutational analysis of specific processes in physiology and development.
2) A rapid means to confirm the identity of a cloned gene. This reverse genetics technology was used to confirm the identity of a candidate clone for the maize an1 gene (Bensen et al., Plant Cell 7; in press). The original clone was isolated by traditional transposon tagging: a screen for dwarf mutants in Mu F2 families, co-segregation analysis of Mu elements with the mutant phenotype, isolation of flanking DNA, recovery of a wild-type clone from a cDNA and/or genomic library. With only a single transposon-tagged allele for this gene, the necessary confirmation (that the cloned sequence was indeed an1) was potentially arduous. However, a reverse genetics screen, using PCR primers specific for the candidate clone and the Mu-TIR primer, uncovered several new Mu alleles of the cloned sequence. Analysis of the F2 progeny for each new allele revealed the expected dwarf phenotype. The clone was thus confirmed as an1; the technology made this type of confirmation possible in weeks, rather than the months or even years necessary to tag additional mutants, or to screen for revertant progeny from the existing mutant.
The main point to be made about this technology is that it is a powerful tool for the study of gene function in plants, specifically maize. Researchers are invited to learn more about this tool, including the opportunity to make use of it. Of greatest importance are the project-specific requirements that must be met before a mutant request is considered formal. This includes development of control alleles and thorough testing of PCR primers. Complex projects, such as those concerning mutagenesis of gene families, naturally require more rigorous planning. For more information, please contact Bob Meeley at the project's e-mail address ([email protected]).
* We wish to thank Vicki Chandler, Michael Freeling, Sarah Hake, Pat Schnable, and Virginia Walbot for donating their "hottest" Mu-active lines.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}