The study of the relationships between transcription and replication of DNA in mitochondria (mtDNA) of different genotypes can provide valuable information on functional organization of the mitochondrial genome of maize. Possible relationships between the transcription and replication in the cytoplasmic organelles of eukaryotes can be implied from the data obtained with vertebrate mitochondria. It is known that the initiation of transcription of the L-strand and the transcriptional activation of the H-strand replication originate from the same DNA site (Clayton, Annu. Rev. Cell. Biol. 7:453-478, 1991). RNA-polymerase has been found in yeast mitochondria, that is likely to provide the synthesis of RNA primer in order to replicate mtDNA (Christianson and Rabinowitz, J. Biol. Chem. 258:14025-14033, 1983). Besides, there is no evidence on the relationship between activities of RNA- and DNA-synthesizing systems in mitochondria of different corn genotypes. Therefore, the purpose of this work was to study the kinetics of the synthesis of RNA and DNA in the isolated mitochondria of different inbreds and hybrids of maize.
In our experiments, the mitochondria used were immediately isolated from the coleoptiles of 3-day-old etiolated seedlings of inbred lines: GK26zakT, W64A, B84zakT, Mo17TV, B73zakT and hybrids: BEKKE LLO, Dneprovsky 320 MV, Odessky 44, Kollectivny 244MV, Rossijsky 1, VIR 46MV, Krasnodarsky 362ATV, Krasnodarsky 303 ATV and Zherebkovsky 86MV. The mitochondria were isolated by a standard method of differential centrifugation. Protein was determined by the Lowry method. DNA synthesis was measured in mitochondria according to the method of Schegget and Borst (Biochim. Biophys. Acta 95:235-248, 1971) with the use of 32P-dATP (specific radioactivity was > 111 Pbq mol-1) or 3H-dATP (980 Tbq mol-1). The conditions for measuring RNA synthesis in mitochondria were essentially similar to those described elsewhere (Carlson et al., Curr. Genet. 11:151, 1986). The kinetics of the RNA synthesis was registered by 32P-UTP (specific activity was > 74 Pbq mol-1) or 3H-UTP (specific activity was 550 Tbq mol-1). The kinetic data were obtained from at least 3-4 experiments.
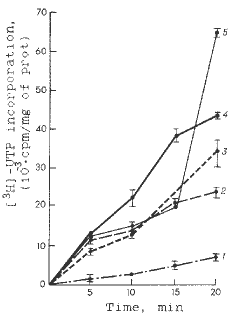
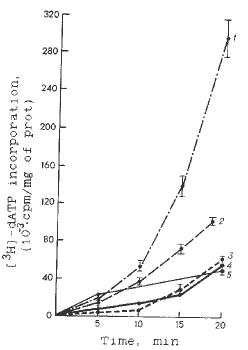
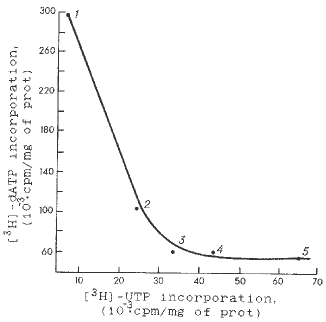
The kinetics of the RNA and DNA syntheses was recorded in the mitochondria of five maize lines within 20 min intervals, and showed that each inbred line had its particular relationships to the activities of the template process of nucleic acid synthesis. Figs. 1 and 2 show low DNA synthesis in mitochondria of maize lines B73zakT, Mo17TV and B84zakT, which possess a high transcriptional activity of mtDNA. Moreover, the mitochondria of GK26zakT lines, which synthesize mtRNA at a much lower rate, show a greater DNA-synthesizing rate. Only line W64A is somewhere in the intermediate position according to the rate of the mitochondrial RNA and DNA syntheses. Thus, we can conclude that in the genetic system of mitochondria of the maize genotypes studied there are reciprocal relations between the rate of RNA and DNA syntheses. In each line, this relation can be followed from the apparent prevalence (except for the W64A line) of one of the two types of template activity of mtDNA. Figure 3 shows the relationships between the characteristics of the mitochondrial systems of nucleic acid syntheses in all maize lines studied. A negative correlation between the activities of these systems can be seen. The coefficient of correlation calculated by the standard formula is -0.99.
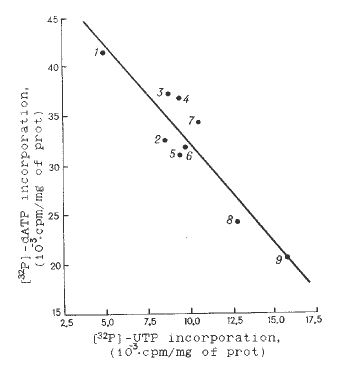
A negative correlation between activities of mtRNA and mtDNA syntheses in organello (coefficient of correlation is -0.92) was also revealed for nucleic acids synthesis in mitochondria of corn hybrids (Figs. 3, 4).
The molecular mechanism responsible for genotype-specific relations of the activities of RNA- and DNA-synthesizing systems of maize mitochondria is still unknown. Neither is its biological significance in the genetic system of these organelles and the cell as a whole. However, it can be assumed that the reciprocal relationships between the synthesis of the two classes of the main genetic macromolecules in mitochondria depend on the size of the corresponding pools of nucleic acid precursors (ribonucleoside triphosphates and desoxyribonucleoside triphosphates), which are formed involving ribonucleotide reductase (EC 1.17.4.1). This enzyme catalyzes the direct recovery of ribonucleoside di- or triphosphates into corresponding desoxy derivatives and provides the DNA synthesis with the necessary pool of deoxyribonucleoside triphosphates (Reichard and Ehrenberg, Science 221:514-519, 1983). In this case, there are favorable conditions for a highly efficient synthesis of mtDNA in genotypes with a high activity of mitochondrial ribonucleotide reductase. Conversely, in genotypes with a low mitochondrial ribonucleotide reductase activity a large-sized pool of ribonucleoside triphosphates contributes to a high activity of the synthesis of the mitochondrial RNA.
In analyzing the results of the present study, we should bear in mind that the prevalence of a single kind of template activity of DNA in maize mitochondria is somehow related to inter-genotype differences, both in the degree of polyploidy of the mitochondrial genome and in the total content of mitochondria in the cell.
Fig. 1. Kinetics of RNA synthesis in isolated mitochondria of different maize lines. 1, GK26; 2, W64A; 3, B94zakT; 4, Mo17TB; 5, B73zakT.
Fig. 2. Kinetics of DNA synthesis in isolated mitochondria of maize lines. Designations are as in Fig. 1.
Fig. 3. Dependence between RNA and DNA syntheses in mitochondria of different maize lines. The organelles were exposed to labeled precursors for 20 min. Designations are as in Fig. 1.
Fig.
4. Dependence between RNA and DNA syntheses in mitochondria of different
maize hybrids. 1, BEKKE LLO; 2, Kollektivny 244MV; 3, Krasnodarsky 303ATV;
4, Zherebkovsky 86MV; 5, Odessky 44; 6, Rossijsky 1; 7, Dneprovsky 320
MV; 8, VIR 46MV; 9, Krasnodarsky 362ATV.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}