The synthesis of two low molecular weight heat shock proteins correlates
with the growth rate of maize lines at enhanced temperature
--G.Borovskii, V.Voinikov, L.Voronova
The response of plants to the influence of high temperature is closely related to the synthesis of specific proteins called heat shock proteins (HSPs) (Lindquist and Craig, Annu. Rev. Genet. 22:631-677, 1988; Vierling, Annu. Rev. Plant Physiol. Plant Mol. Biol. 42:579-620, 1991, for review). The adaptive role of HSP synthesis is well known and the physiological functions of a number of proteins are fairly well studied. The role of low molecular weight (LMW) HSPs of plants remains less studied at present. The diversity of this group of proteins makes it particularly difficult to determine their functions.
Seedlings of inbred lines were used as a model system to give a preliminary determination of LMW HSPs in order to examine them at length. The growth response of young plants to high temperature indicates the ability of the line to adapt to experimental conditions. The investigations performed showed a relation between the synthesis of two LMW HSPs and the ability of the line to enhance the rate of growth at high temperature.
The following inbred lines were used in the study: A34, A334, A166, L, Ma21, H513, S54, SV56, Oh43, HY WS, W32, V158, A344, Ma23C, A340, A295, A347, A375, A641, PLS61 and W834 (kindly provided by Krasnodar Research Institute of Agriculture, Russia). The adaptive ability of the seedlings and the set of low molecular heat shock proteins were studied. Three-day-old seedlings were transferred to a growth chamber on a medium containing Knopp nutrient solution. The first group of plants was kept at 28 C and the second one at 38 C. The relative humidity was 60%. The shoot length was measured in 30 plants of each group every day starting from the fifth day of germination. The measurements were performed up to the stage of the appearance of the fifth leaf, i.e. 13-15 days from the emergence of seedlings. The curve of changes in the above-ground part of the seedlings in both groups was of an S-shaped form during the experiment. The coefficients were calculated in the equation of linear regression for curves showing the change in the length of the above-ground part of the seedlings. In this case, the coefficient of regression is the average growth rate of plants of each line at 28 C or 38 C. To calculate the coefficients of regression (R1 at 28 C and R2 at 38 C), the middle part of the curve, an almost linear one (from the 6th to the 11th day of germination), was used. The ratio of the coefficient of regression at 38 C to that at 28 C shows changes in the growth rate under the effect of high temperature and the ability of plants to adapt to it. This ratio was called the coefficient of thermo-adaptation (Ct). Average coefficients of regression and coefficients of variation were calculated from the sampling of coefficients of regression at 28 C and 38 C in the lines studied.
Table 1. The growth rate of varied maize lines at control (28 C) and
high (38 C) temperature, indicated by regression coefficients (R1
and R2) and thermoadaptation coefficient (Ct) which
is R1/R2.
| 28C | 38C | ||
| Line | R1 | R2 | Ct |
| A334 | 29.45 ± 2.01 | 38.55 ± 2.55 | 1.31 |
| A347 | 27.9 ± 2.27 | 34.69 ± 2.40 | 1.24 |
| V158 | 25.09 ± 0.97 | 30.14 ± 0.99 | 1.20 |
| A34 | 21.1 ± 1.10 | 25.0 ± 0.67 | 1.18 |
| L | 22.77 ± 1.55 | 26.4 ± 0.66 | 1.16 |
| SV56 | 28.1 ± 1.36 | 31.85 ± 1.44 | 1.13 |
| W32 | 24.46 ± 0.06 | 27.2 ± 0.84 | 1.11 |
| HyWS | 22.1 ± 0.92 | 22.4 ± 0.80 | 1.08 |
| A641 | 29.43 ± 1.78 | 31.51 ± 1.04 | 1.07 |
| Oh43 | 20.99 ± 1.26 | 22.4 ± 0.89 | 1.07 |
| A166 | 30.29 ± 2.60 | 31.9 ± 1.04 | 1.05 |
| A344 | 26.8 ± 1.30 | 26.47 ± 0.78 | 0.99 |
| S54 | 30.2 ± 1.65 | 29.4 ± 1.19 | 0.97 |
| PLS61 | 34.15 ± 1.30 | 32.9 ± 0.95 | 0.96 |
| A295 | 27.2 ± 1.10 | 25.6 ± 1.00 | 0.94 |
| Ma23C | 26.89 ± 0.90 | 24.88 ± 1.32 | 0.93 |
| A375 | 28.29 ± 1.19 | 25.04 ± 0.64 | 0.89 |
| W834 | 22.75 ± 1.03 | 19.5 ± 0.64 | 0.86 |
| A340 | 27.2 ± 1.57 | 23.2 ± 1.02 | 0.85 |
| H513 | 29.29 ± 1.43 | 24.77 ± 1.16 | 0.85 |
| Ma21 | 26.8 ± 1.70 | 20.6 ± 1.60 | 0.77 |
The temperature of 38 C appears to be critical for seedlings. At hyperthermia the growth rate of the seedlings increases, decreases, or remains unchanged (Table 1). Thus, lines can be distinguished by varying responses (according to the growth rate) to the high temperature effect. The average value of the coefficient of regression did not change greatly at the high temperature: 26.73 at 28 C and 27.35 at 38 C. However, the coefficient of variation increased from 12.76 at 28 C to 17.95 at 38 C. This increase resulted from varying changes in the growth rate in different genotypes under conditions of high temperatures. The ability of the seedlings to grow at 38 C, and hence the adaptive potentials of the given genotype are expressed by the coefficient of thermo-adaptation, Ct (R2/R1 ratio). This coefficient varied from 0.77 (Ma21 line) to 1.31 (A334 line).
To analyze the set of LMW HSPs, eight lines were selected. Three of them showed various degrees of inhibition of growth at high temperature: Ma21 (Ct=0.77), H513 (Ct=0.85) and S54 (Ct=0.97). In other lines the Ct increased in different degrees: A166 (Ct=1.05), SV56 (Ct=1.13), L (Ct=1.16), A34 (Ct=1.18) and A334 (Ct=1.31). The 3-day-old etiolated seedlings of selected lines were incubated in the presence of 35S-methionine for three hrs at 28 C (control) and 41 C (heat shock). The labeled proteins were used in electrophoresis according to Laemmli (Nature 227:680-685, 1970). Radiographs of the gels were analyzed using a densitometer.
All lines were found to synthesize the heat shock proteins at hyperthermia, though their number and the intensity of the synthesis varied. Table 2 includes all polypeptides with molecular weight (MW) from 9 to 32 kDa if the synthesis of this polypeptide was enhanced at increased temperature in at least a single line studied.
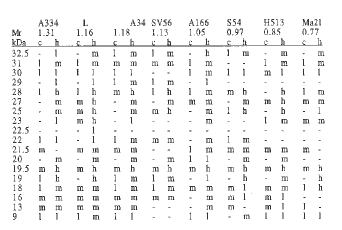
Table 2. The synthesis of low molecular weight heat shock proteins by varied maize lines (Ct is given just beneath each maize line). The seedlings were incubated at 27 C (c) and 41 C (h) for 3 h. The labeled proteins were separated by SDS-PAGE. Radiographs of the gels were analyzed on a densitometer. The synthesis was on high (h), medium (m) and low (l) level, or not detected (-).
The diversity of the LMW HSPs observed is rather high. Eighteen groups of polypeptides with varying MW were obtained. To facilitate the analysis, the polypeptides were separated into major, i.e. being actively synthesized, and minor ones with poor label incorporation. The major groups were composed of HSPs with MW 32.5, 31, 28, 27, 25, 19.5, 19 and 18 kDa. Other polypeptides belong to the minor groups.
A number of major HSPs with MW 32.5, 28, 27, 25, 19.5 and 19 kDa were synthesized in all lines studied. Only the S54 line failed to synthesize two major HSPs with MW 31 and 18 kDa (Table 2). Looking for a possible relation between the coefficient of thermo-adaptation and the HSP synthesis, attention was paid to the minor HSPs with MW 29 and 16 kDa. The 29 HSP was synthesized only in lines growing faster at the high temperature than at the control temperature. The protein with MW 16 kDa was synthesized in lines H513 and S54 at the control temperature but was greatly inhibited by heat shock. This protein was completely absent in the Ma21 line with the lowest Ct. The synthesis of this polypeptide increased or was maintained at the same level only in lines with the coefficient of thermo-adaptation greater than one unit. The data suggest a relation between the synthesis of HSPs 29 and 16 and the adaptation processes in young plants at high temperature.
The mechanism of the effect of the HSPs 29 and 16 on the capacity of maize to grow intensively at enhanced temperatures is obscure. It may be related to the effect of these proteins on the effective function of the mitochondria at high temperatures. The association of some LMW HSPs with seedling mitochondria has been shown upon heat shock in vivo. In this case the HSPs 29 and 16 were associated with the outer membrane of these organelles (Borovskii and Vojnikov, Russian J. Plant Physiol. 40:596-598, 1993). Furthermore, it is known that the association of LMW HSPs with mitochondria contributes to the oxidative phosphorylation abilities of isolated mitochondria in vitro (Chou et al., Plant Physiol. 89:617-621, 1989; Vojnikov et al., Plant Physiol. (Life Sci. Adv.) 8:1-4, 1989). Recently the strong thermo-stabilization influence of plant HSPs with MW 15-18 kDa on soluble and membrane associated proteins has been reported (Jinn et al., Plant Cell Physiol. 34: 1055-1062, 1993). The molecular mechanism of these proteins' action may be related to chaperone, since a chaperone function has been demonstrated in vitro for a mammalian LMW HSPs (Jacob et al., J. Biol. Chem. 268:1517-1520, 1993).
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}