Localization of the Pex1 protein to the pollen tube wall
-- Amy Rubinstein, Anne Broadwater and Patricia Bedinger
Very little is known about the genes required for pollination in self-compatible species, such as maize, or about the mechanism of the rapid growth of the maize pollen tube. As a first step toward identifying some of these genes, our lab isolated several genes expressed specifically in pollen. Two of the clones isolated contained a very distinctive repeat motif: ser-pro4. Prolines found in this motif are usually hydroxylated. Hydroxyproline is an unusual amino acid that occurs in only a few groups of extracellular matrix proteins, including collagen and elastin in animals and hydroxyproline rich glycoproteins (HRGPs) in plants. One major class of HRGPs, the extensins, are characterized by the ser-pro4 motif that we found in our clones. Thus, we named the genes Pex1 and Pex2 (pollen, extensin-like).
Finding genes like these in pollen was an exciting discovery for a number of reasons. First, although extensin-like genes have been found to be specifically expressed in the pistil, none had been found in pollen. These are the first pollen-specific genes to be found with the canonical extensin repeat motif. Secondly, the discovery of these genes also marked the first time a classical extensin repeat motif had been found in any maize gene.
We isolated and sequenced a complete genomic clone representing Pex1. We discovered that the gene actually contained two distinct domains: one contained the ser-pro4 repeats and was presumed to have a rod-like structure. The other domain, at the N terminus, contained no repeated motifs. This structure was reminiscent of the sexual agglutinins of Chlamydomonas, which mediate the recognition between the plus and minus mating types of Chlamydomonas. We would now like to investigate the possibility that the pollen Pex proteins interact with a partner molecule in silk in a manner analogous to mating in Chlamydomonas.
Southern analysis confirmed that there are two copies of this gene in the maize genome. From Northern analysis, we determined that the Pex genes are expressed specifically in pollen. In addition, we found that the Pex genes are not expressed in early stages of pollen development and are present at a low level in starch-filled immature pollen. Only upon pollen maturation were high levels of Pex messages detected. This work, including the cloning and characterization of the Pex genes, has been accepted for publication in the Proceedings of the National Academy of Sciences, USA.
To more directly study the protein encoded by Pex1, and to provide clues about its function, a fusion protein was constructed using a piece of the Pex gene and the trpE gene of E. coli. Antibodies raised against this fusion protein recognized two proteins in pollen extracts just over 200 kD in size, as well as larger proteins in the stacking gel. In agreement with Northern analysis, these proteins were not present in seedling, endosperm, ear, silk, root, leaf or wounded leaf extracts. We also found no Pex protein in meiotic tassel or young microspore extracts. In starch-filled immature pollen, the Pex antibodies recognized the >200 kD proteins, but did not recognize any proteins in the stacking gel. Because deglycosylation with trifluoromethanesulfonic acid (TFMS) removed the proteins in the stacking gel, we are hypothesizing that they represent the glycosylated form of the Pex proteins. Our results suggest that the glycosylation of the Pex proteins occurs during very late stages of pollen development.
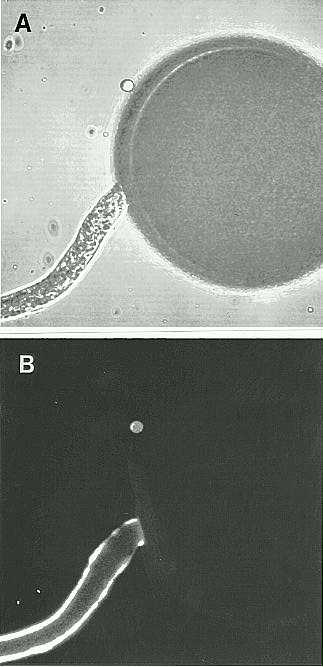
We have also used the Pex antiserum in immunolocalization experiments with in vitro germinated pollen. Utilizing a FITC-labeled secondary antibody and with the aid of a confocal microscope, we were able to determine that the Pex proteins are localized to the pollen tube wall. This result is consistent with a function for the Pex proteins during the process of pollination.
In addition, we carried out several experiments to determine the nature of the association of the Pex proteins with the pollen tube wall. The extensins, for example, are known to be covalently crosslinked to the walls of vegetative cells. To test for such tight association, we treated pollen tubes with 1% SDS, 1M NaCl and 8M LiCl. None of these agents eliminated the Pex signal. These results indicate that the Pex protein is tightly bound (possibly covalently) to the pollen tube wall. This is consistent with the properties of known extensins, which cannot be extracted from cell walls with SDS or high salt. Treatment of the pollen tubes with Proteinase K caused the Pex signal to disappear almost entirely. This indicates that the substance being recognized by the antibody is indeed a protein.
We envision three possible functions for the protein product of the Pex genes. It could be a structural component of either the pollen or pollen tube walls, necessary for providing structural support during the extremely rapid growth of the pollen tube. It may be involved in the initial recognition between pollen and silk at the point of pollen tube germination, or it may mediate cell-cell signaling during pollen tube growth. It is also possible that the Pex proteins are involved in all of these processes.
Figure
1. Localization of Pex1 to the pollen tube wall with the Pex1 antibody
and a fluorescent secondary antibody using a confocal light microscope.
A. transmitted image B. fluorescent image.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}