Hunter and Dalbey (Am. J. Bot. 24:492-494, 1937) studied histological structures in maize looking for accurate indicators of the relative degree of stalk breakage in the field. Since Sibale et al. (Maydica 37:111-114, 1992) compared the mechanical force gage (manual), vs. a modified-digital force gage (electronic) rind penetrometer, studies at the University of Missouri (Masole, M. Sc. thesis, 1993; Chumo, Ph. D. thesis, 1993; and Alsirt, M. Sc. thesis, 1993), have shown that the Missouri-modified electronic rind penetrometer was easier, more accurate, and more consistent in ranking genotypes (CVs were 7.5, 10.4, and 14.8%, respectively, for the three thesis studies). The benefit of using the rind penetrometer is greater because it is a non-destructive method; therefore, selection can be done before anthesis and the selected plants can then be intermated in the same season. This allows for completion of one cycle of selection per season in a recurrent selection program.
In maize, stiff-stalked genotypes have been characterized by thick bundle sheaths, thick walled, sub-epidermal sclerenchyma, and more angular, close-fitting cells. Weak stalks were characterized by thin walls, narrow bundle sheaths, and large intercellular spaces. Morphological characters that showed a consistent relationship to stalk strength were low bundle number mm-2 in the rind, high percentage of sheath bundle-1, large stalk diameter, and wide lignified zone (Magee, Iowa State Coll. J. Sci. 22:257-268, 1948). The extent of lignification beyond the rind and the number of vascular bundles within this lignified area were correlated with lodging resistance in maize (McRostie and MacLachlan, I. Sci. Agr. 22:307-313, 1941). Zuber and Grogan (Crop Sci. 1:378-380, 1961) found a significant correlation between mechanical strength and rind thickness. The number of vascular bundles was lower, the thickness of the sclerified parenchyma was greater, and thickness of bundle sheaths was greater in strong-stalked as compared to weak-stalked genotypes (Kálmán et al., Acta Biol. Szeged 21:[1-4],35-40, 1975).
The objective of this project was to identify changes in stalk anatomy
in response to bi-directional selection for rind penetrometer resistance.
Stalk sections of a representative plant from each extreme of selection,
C6 high (RPR=19.9 load-kg) and C6 low (RPR=2.9 load-kg), were obtained
from the middle portion of the internode used for selection for RPR. Stalks
were cut into about 5-mm thick cross sections and fixed in a formaldehyde-alcohol-acetic
acid (FAA) solution (50 mL 95% EtOH, 5 mL glacial acetic acid, 10 mL 37-40%
formaldehyde, and 35 mL dH2O). Sections were fixed for 48 h, taken out
of FAA, and washed in 70% and 50% EtOH for 1 h each. After washing, sections
were put in Hepes (N-[2-Hydroxyethyl] piperazine-N'-[2-ethanesulfonic acid],
Sigma Co., St. Louis, MO) buffer for 3 to 4 h, changing the buffer twice
during this time, and leaving the samples in Hepes buffer for long-term
storage. Hepes buffer keeps samples at physiological pH (7.2 to 8.2), better
preserving microstructures. Attempts were made to obtain thin sections
(10 to 20 mm) using a Cryo-Stat microtome (Jung Frigocut 2800N, Leica Instruments
GmbH, Nussloch, Germany). The Cryo-Stat microtome requires small samples
mounted in a plastic polymer Histo-Prep (2-phenoxyethanol, polyoxypropylene-polyoxyethylene
block copolymer, Fisher Sci., Fair Lawn, NJ) embedding media and frozen
at temperatures below 0?C. Cryo-sectioning is a fast procedure designed
to obtain numerous sections in a relatively short period of time. Unfortunately,
the different consistency of tissues, specifically those from C6 high,
made it impossible to obtain tear-free sections. Alternatively, samples
embedded in paraffin and sectioned at 10 Am with a manual microtome were
successfully obtained for both cycles. To eliminate paraffin from sections
once they were mounted on the glass microscope slide, sections were placed
in xylene for 10 min and then rehydrated by placing them in two washes
of 100% EtOH and 95% EtOH, for 10 min twice each, and once each in 70%
dH2O and Hepes. Non-stained preparations were used for observations under
the confocal microscope. To identify patterns of lignin accumulation, a
stain specific for lignin developed by Conn (Biological Stains, 4th ed.,
The Williams and Wilkins Co., Baltimore, MD., 1953) was used. The applicability
of this stain is for general plant tissues, especially meristematic tissue,
and it is designed to show chromatin, spindles, cellulose, and lignified
walls. The staining procedure involved: (i) removal of paraffin from sections,
(ii) staining for 30 to 50 min in 1% aqueous safranin-o, (iii) rinsing
in dH2O, (iv) counterstaining with 0.2% fast green in EtOH for 1 to 1.5
min, and (v) rinsing in absolute EtOH. Stained sections were permanently
fixed on the slide by saturating with polymer Mowial (Calbiochem Corp,
La Jolla, CA) and covered with a glass cover.
Fixed slides were observed under a MRC-600 (BIO RAD, Cambridge, MA)
laser scanning confocal microscope and digitized with a Nikon (Optiphot,
Nikon Corp., Tokyo, Japan) video camera at the University of Missouri-Columbia
Molecular Cytology Core facility. Taking advantage of tissue autofluorescence,
images from non-stained preparations of C6 high and C6 low were captured
in a Macintosh Quadra (Model 840 AV, Apple Computer, Inc., Cupertino, CA)
computer and dye-sublimation-printed on thermal graphic paper. Preparations
stained with safranin-o were observed under a bright-field light microscope
and images were captured by an Image-1/AT (Fryer Co., St. Louis, MO) image
analysis system. Final printing was also done on a dye-sublimation printer
(Codonics, Inc., Akron, OH).
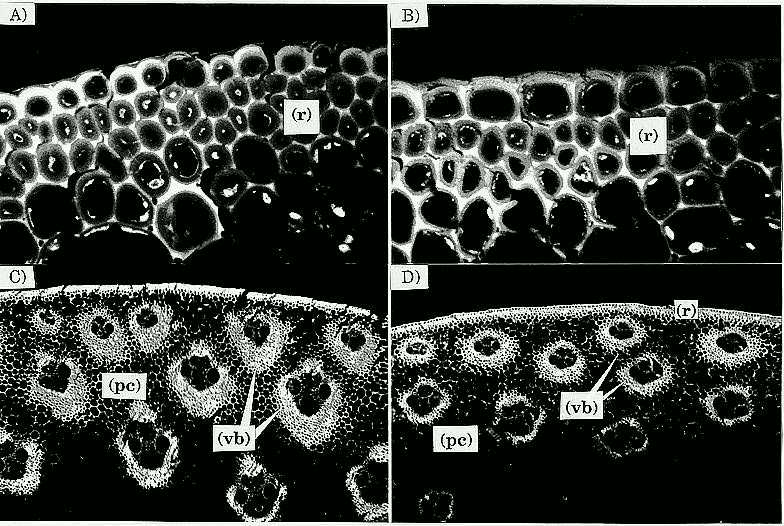
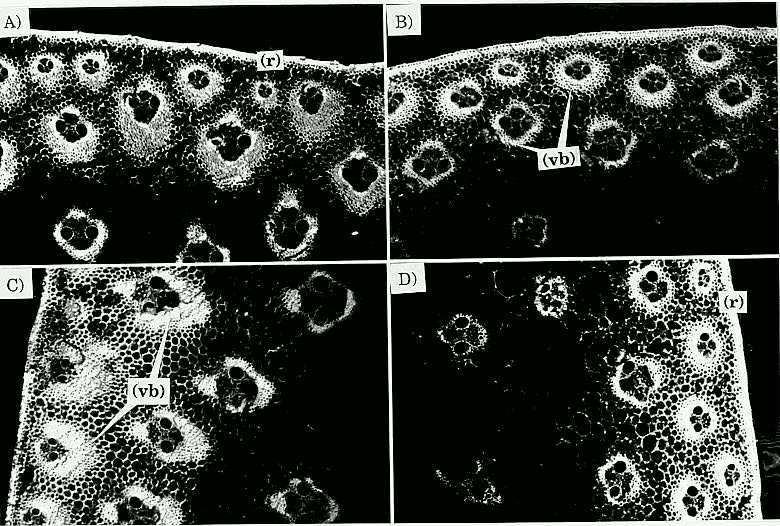
Sectioning the C6 low stalk material was easier than C6 high, apparently because stalks from C6 low have more similar rind and pith tissue consistency, whereas the C6 high stalks have two different tissue consistencies with the rind being much tougher than the pith. The main differences between sections of C6 low vs. C6 high as observed under the confocal microscope were that in C6 high: i) the rind, mainly composed of parenchyma cells, was denser, and perhaps had one or two extra layers of cells; ii) the parenchyma cell walls were thicker; iii) a more angular shape of vascular bundle sheath region, defined by a greater density of cell layers forming the sheath; and iv) the inward portion of the rind vascular bundle sheaths (sclerenchyma cells) was larger (Fig. 1). Stalks from C6 high have a parenchyma cell region (in which the rind vascular bundles are contained) that extends further towards the center of the stalk than in C6 low stalks (Fig. 2). These findings agree with those previously reported by Magee (1948), Kálmán et al. (1975), Berzonsky et al. (Crop Sci. 26:482-488, 1986), although the mechanisms that produced such changes may be different. No apparent difference in vascular bundle numbers between C6 low and C6 high was observed, which contrasts with the findings in MoSQA and MoSQB, whose vascular bundle numbers were greater in the high than in the low selections (Chang et al., Crop Sci. 16:621-625, 1976; and Berzonsky et al., 1986). However, the density of vascular bundles area-1 was greater in the C6 high stalks because the stalk diameter was smaller in C6 high than in C6 low.
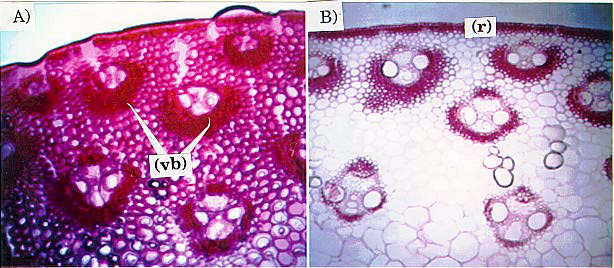
In an attempt to identify differences in lignification, cross-sections
of C6 low and C6 high were stained for lignin using o-safranin and fast
green. This particular stain is designed to show
cellulose and lignified structures in a red color. Histological sections
of the internode used in selection for RPR showed differences in accumulation
of lignin around the vascular bundles of plants from the extremes of selection
(C6 low vs. C6 high; Fig.
3). It is likely that the differential in lignin accumulation is a
correlated response to selection, rather than a direct response. The staining
patterns agreed with vascular bundle sheath patterns shown in Figs. 1 and
2. Rind vascular bundle sheaths in C6 high were more heavily stained than
those from C6 low, indicating a greater lignification of cell walls in
the C6 high stalks.
Characterization of C6 high agrees with results reported by Chang et al. (1976) and McRostie and MacLachlan (1941). Chang et al. (1976) indicated that MoSQB, selected for high stalk crushing strength, had more heavily lignified vascular bundles, whereas McRostie and MacLachlan indicated that lodging-resistant lines had considerably more lignification within the rind than lodging-susceptible lines.
A possible mechanism by which plants selected for high RPR achieve stalk strength is through the condensation of the rind parenchyma cell layers and rind vascular bundle sheath cell layers, and thickening of the cell wall, along with lignification of such structures. The condensation of stalk structural components may be reflected in findings by Masole (1993), who showed increase of rind thickness and reduction of stalk diameter in response to selection for high RPR.
Fig. 1 (following page, top) Confocal micrographs of stalk cross sections of C6HRP and C6 LRP. Compare: A) Rind of C6 HRP (60X); B) Rind of C6 LRP (60X); note more layers of rind parenchyma cells and thicker interlumen region in A as compared to B; C) C6 HRP (4X); and D) C6 LRP (4X); note more angular shape and larger inward portion of vascular bundle sheaths of B as compared to C. Rind = (r); parenchyma cell region = (pc); and vascular bundle-sheaths = (vb).
Fig. 2 (following page, bottom). Confocal micrograph comparison of stalk cross sections of C6 HRP (A and C) and C6 LRP (B and D) (4X). Note the more angular shape and larger inward portion of vascular bundle sheaths of A and C compared to B and D. Also note that the parenchyma cell region (pc) in C extends further inward. Rind = (r); parenchyma cell region = (pc); and vascular bundle-sheaths = (vb).
Fig.
3. Light micrographs comparing staining for lignin in stalk crosssections
of C6 HRP (A) and C6 LRP (B) (4X). Rind (r), parenchyma cell region (pc)
and vascular bundle-sheaths (vb) are shown. Note a more general retaining
of the stain and more heavily stained vascular bundle sheaths in A as compared
to B. Stain also concentrated on the vascular bundle-sheaths shown in Figs.
1 and 2.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}