Our present model of the Ac transposition process proposes that the transposase (TPase) protein binds as an oligomer to internal DNA sites near the termini of the transposon. We assume that the TPase is responsible for bringing the two Ac ends and possibly also the integration site in close proximity during transposition, forming a "transposom".
Two previous observations indicate the occurrence of protein-protein interactions between Ac TPase units. 1) By in situ immunofluorescence staining of transfected Petunia protoplasts large nuclear TPase aggregates were detected. The observation that certain amino acid substitutions in the TPase protein appear to be aggregation-deficient indicates that aggregation is an intrinsic property of the TPase. 2) A number of transposition- and DNA binding-deficient TPase mutants act as dominant inhibitors of transposition when coexpressed with a functional TPase protein.
We have begun using the yeast "two hybrid system" (Fields and Sternglanz,
Trends Genet. 10:286-292, 1994) to localize the TPase protein-protein interaction
domains. The two hybrid system is based on transcription activation of
the lacZ gene by binding of the GAL4 protein to the upstream activating
sequences. The N-terminal, DNA-binding domain of GAL4 may be separated
from the C-terminal transcription activation domain. By fusing these domains
separately with other proteins, ß-galactosidase expression is activated
if the fusion proteins interact.
The full length Ac TPase protein (TPase1-807) and the transpositionally
active deletion derivative TPase103-807 were fused to each GAL4 domain
of the standard two hybrid vectors pGAD424 and pGBT9, and transformed into
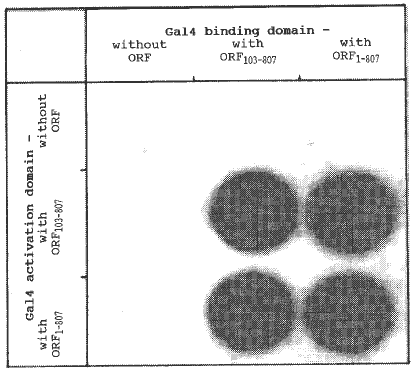
yeast cells. Our results demonstrate that the wildtype Ac TPase
protein can directly interact with itself as well as with the truncated
TPase starting at codon 103 (Figure 1).
We will continue these experiments with deleted Ac TPase segments to localize the interactive domain(s). Although the results are not yet complete we are confident to have identified a small C-terminal TPase fragment which can still interact with the TPase protein starting at codon 103.
Figure
1. The figure shows the ß-galactosidase activities of yeast cells
transformed with different combinations of the two hybrid plasmids. Negative
controls are tests in which the TPase open-reading frame (ORF) is not present
in both plasmids. All cells were grown in liquid suspension culture and
aliquots were filtered through a defined area of a nylon membrane. Subsequently
the cells on the filter were cracked in liquid nitrogen and incubated with
X-GAL at 37 C.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}