In Drosophila and Caenorhabditis elegans, site-selected transposon mutagenesis by PCR screening has proven to be a powerful method for isolating insertions and deletions in known genes to ascertain their function via gene knockouts (Zwaal, et al., Proc. Natl. Acad. Sci. USA 90:7431-7435, 1993; Kaiser and Goodwin, Proc. Natl. Acad. Sci. USA 87:1686-1690, 1990; Ballinger and Benzer, Proc. Natl. Acad. Sci. USA 86:9402-9406, 1989). We have developed a similar method in maize to rapidly identify new insertions and deletions into known genes using Robertson's Mutator. The hcf106-mum1 mutation in maize is caused by the insertion of a Mu1 transposable element into the promoter region of a gene required for chloroplast development (Martienssen et al., EMBO J. 8:1633-1640, 1989; Barkan and Martienssen, PNAS 88:3502-3506, 1991). The hcf106 mutant phenotype depends on transposon activity, and as a result, the phenotype is leaky and difficult to maintain in new genetic backgrounds (Martienssen et al., Genes Dev. 4:331-343, 1990). In order to obtain stable alleles, we used a PCR screening strategy to select plants that had suffered a deletion flanking the Mu1 element at the hcf106 locus. 1500 seedlings from the cross hcf106/+ x +/+ were germinated in the greenhouse in 2-dimensional grids to facilitate pooling. Mu activity was monitored using a second independent mutation (Les28; Martienssen and Baron, Genetics 136:1157-1170, 1994), and calculations showed that the 1500 seedlings represented a collection of about 350 Mu active hcf106 gametes, 350 Mu-active + gametes and 800 gametes that had lost Mutator activity.
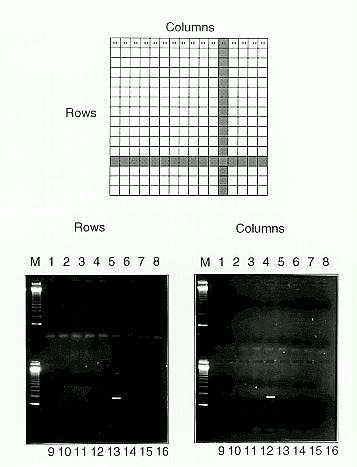
DNA was prepared from pools of 32 seedlings from each row (x) and column (y) of the grids. The DNA was digested with an enzyme that cuts uniquely in the hcf106 gene at the initiator ATG codon. DNA from the hcf106 locus was then amplified using a primer from the promoter and a primer from the second exon of the hcf106 gene. Digestion between the primers ensured that only alleles that had lost the initiator ATG codon via deletion would be amplified in subsequent PCR. Three seedlings were identified by cross-referencing x and y pools that had PCR products of the predicted size (Figure 1). DNA was made from these 3 seedlings individually, and they were found to have suffered a 244bp deletion, insertion of a Mu1 and insertion of a dMuDR element into the second intron of the progenitor, hcf106-mum1, and the normal Hcf106 alleles, respectively (Figure 2). Insertions between the ATG and the second exon were recovered because they allowed amplification with the primers used. Insertions and deletions affecting the 360bp target region between the ATG and the second primer were recovered at a frequency of 1 in 350 gametes. Outcrossing and self-pollination revealed that two of these new alleles (the deletion and one of the insertions) conferred stable mutant phenotypes when homozygous, even in the absence of Robertson's Mutator activity. In contrast, Mutator activity is required for phenotypic expression of the progenitor allele, hcf106-mum1, in homozygous seedlings (Martienssen et al., Genes Dev. 4:331-343, 1990).
This method provides a powerful way to identify deletion (null) derivative alleles in maize, as well as new insertions in maize genes without phenotypic selection: First, insertions are recovered in a gene of known sequence by PCR screening. In a second generation, deletions can be recovered in screens using the first insertion as the starting point. New alleles are recovered in heterozygotes, so that even if the deletion alleles are lethal, they can still be propagated and studied. Sequencing of the PCR products can be used to precisely define the derivative alleles before their phenotype is known, so that this technique is ideal for studying genes with lethal phenotypes, or those for which only dominant, gain-of-function alleles are known.
Figure 1. PCR pooling strategy for site-selected transposon mutagenesis. Seed were germinated in a 16 x 16 grid, with 2 seedlings at each position in the grid as shown. DNA was extracted from pools of 32 seedlings from each row and each column, and amplified using primers from the Mu1 element and the second exon of the Hcf106 gene. Each lane of each gel was loaded with PCR products from individual rows and columns. In this example, the same derivative allele was amplified in one row and one column, and thus identified by a unique address.
Figure
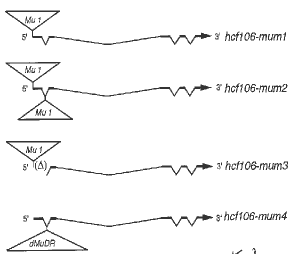
2. Schematic maps of progenitor hcf106-mum1 and derivative hcf106-mum2,
hcf106-mum3,
and hcf106-mum4 alleles. Exon sequences are represented by horizontal
lines, while introns are shown as dips. The deletion in hcf106-mum3
is shown as a triangle.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}