The regulation of C and N supply to developing kernels by the vegetative portions of the plant is a subject of great interest because it is responsible for the accumulation of starch, protein and corresponding increase in dry weight of the kernels. Accordingly, in order to clarify the metabolic factors regulating the relative content of starch and protein in the seed we focused our attention on two strains, Illinois High Protein (IHP) and Illinois Low Protein (ILP), which differ greatly in their ability to accumulate proteins and starch in the endosperm.
Studies of IHP and ILP previously reported by our laboratory suggested that the accumulation of endosperm protein is affected by the amount of N substrates supplied to the kernel. We believe additional information about the processes involved in N metabolism will prove useful in elucidating the physiological bases involved in N utilization in maize plants.
Plants of IHP and ILP strains were grown under field conditions at Bergamo with different levels of N fertilizer (0, 75, and 150 Kg/ha N), to study the accumulation and the partitioning of N in different plant parts. At two dates, i.e. silking stage and black layer maturity (BLM), 3 whole plants (total above ground parts) from each of 4 replications from each N level, were separated into leaves, stalks, and ears, which were further divided into husks, cobs, and grains at the second harvest and analyzed for total N content. Plants of the IHP and ILP strains showed a different behavior in response to the increasing rate of N fertilizer application. In particular, it was noted that plants from the IHP strain had a higher N uptake and N redistribution efficiency than plants from the ILP strain. For the IHP strain, measurements show that increasing the rate of N fertilizer application is accompanied by an increase in N concentration in the whole plant at silking and by an increase of N concentration in the kernel at BLM. The ILP strain responded to N application only at the higher level of N application. Data at BLM revealed that for ILP plants the N concentration of the whole plant, with the exception of grain, was similar to the N concentration observed at the silking stage. These results suggested that the ILP kernel phenotype is apparently due to a reduced ability of N relocation from the plant to the developing grains.
A second field experiment was conducted to document the partitioning of 15N applied to IHP and ILP strains, and to follow the pattern of subsequent remobilization of this N during plant development. For each genotype 340 mg of labeled N (as K15NO3, 14.4 atom % excess) was applied to individual plants in the two central rows of each subplot.
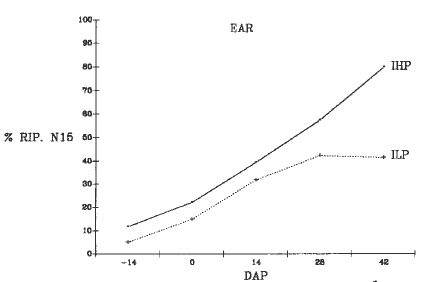
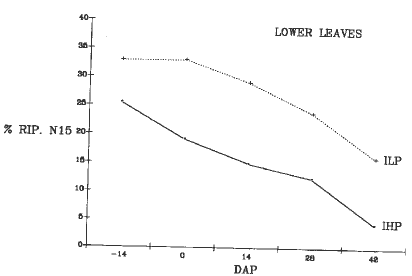
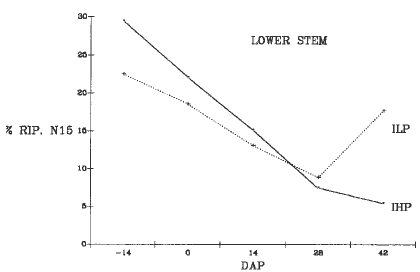
Plants were harvested at 5 stages: 14 days before pollination (-14 DAP), at flowering (0 DAP), 14-28 DAP, and at BLM. The plants were divided into 8 parts; the 15N concentration was determined with a mass spectrometer (Tracer Mass Europa Scientific) connected to the N-analyzer (Carlo Erba 1500-NA). During ear development the IHP strain showed a higher efficiency in relocating the 15N to the ear than the ILP strain (Fig. 1). In the other plant parts during the grain filling period, a greater decrease of 15N concentration was evident in the IHP strain than in ILP, particularly in the lower leaves (Fig. 2) and in the lower stalks (Fig. 3). The ILP strain showed a decrease in the percentage of 15N distribution in the lower leaves until 42 DAP, in the lower stalks until 28 DAP, and a concomitant increase of 15N in the ear until 28 DAP. These results suggest that the ILP phenotype depends on a lower ability of N remobilization during the late stage of grain development.
Figure 1. Change of 15N accumulation in the ear after the labeling period.
Figure 2. Change of 15N accumulation in the lower leaves after the labeling period.
Figure 3. Change of 15N accumulation in the lower stems after the labeling period.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}