Among the many families of maize transposable elements, Mutator (Mu) elements are particularly diverse. While the defective elements of the En(Spm) and Ac/Ds families are mostly deletion derivatives of the respective autonomous element, Mu elements can be divided into subfamilies that have largely unrelated internal sequences. Another unusual feature of the Mu element family is their relatively long (185-514 bp) terminal inverted repeats (TIRs).
The autonomous Mu element, MuDR (Chomet et al., Genetics 129:261-270, 1991; Hershberger et al., PNAS 88:10198-10202,1991; Qin et al., Genetics 129:845-854,1991), is 4942 bp long and encodes at least two convergent transcripts (Benito and Walbot, Maydica, 1994). Six subfamilies of Mu elements, based on similarities of the internal sequences, are currently accepted: Mu1/Mu2, Mu3, Mu4, Mu6/Mu7, Mu8 and MuDR/Mu5 (Bennetzen et al., Crit Rev Plant Sci 12:57-95, 1993).
The sources of the diversity found in the Mu element family have not been fully identified, and different models on how these elements evolved have been proposed (Talbert et al., J Mol Evol 29:28-39, 1989; Bennetzen and Springer, TAG 87:657-667, 1994). At least some internal changes involved acquisition of large segments of DNA from other parts of the genome, making it difficult to determine exactly what events, how many events, or what order of events have occurred. Because the ends of the elements are more conserved than internal sequences, differing primarily by single base substitutions, a relatedness analysis of this element component seemed feasible.
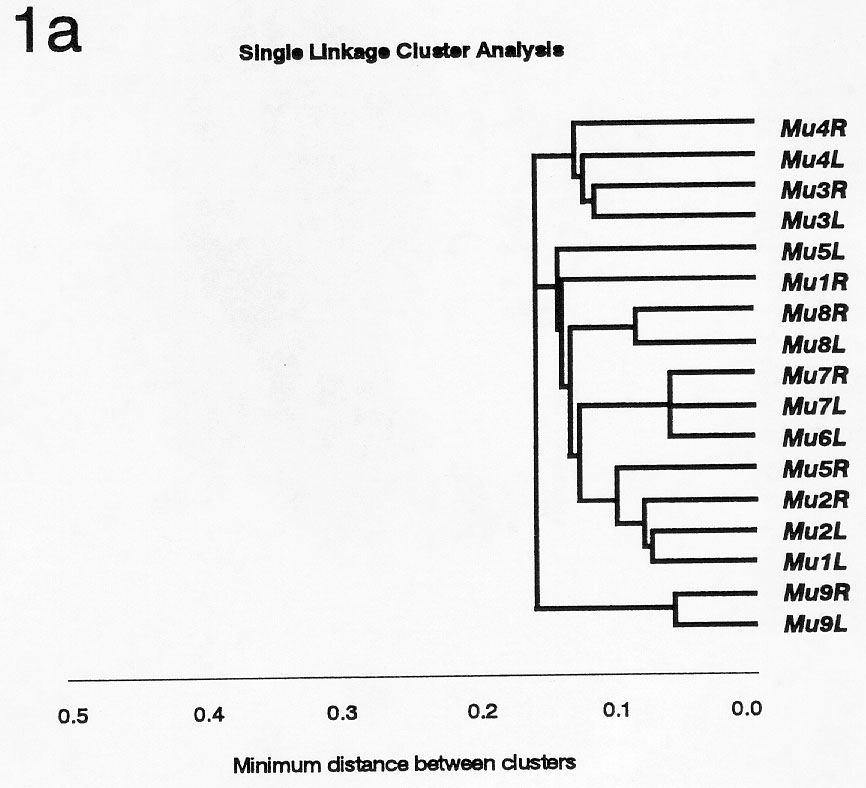
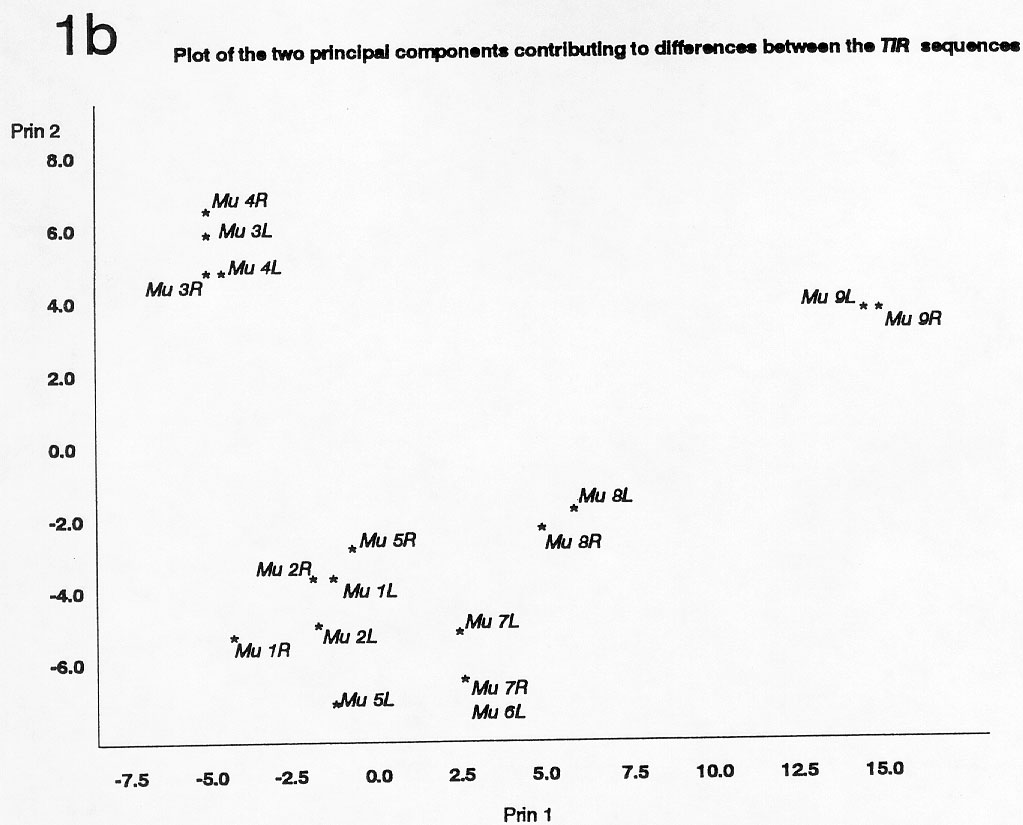
For each of the elements, either the most-terminal 172 bp (Mu6 's left TIR and Mu3 's right TIR) or 171 bp (all other sequenced Mu TIRs) were found to be most conserved between elements, and these sequences were used for comparisons. Cluster and Principal Components analyses of these 171 bp or 172 bp of Mu TIRs are shown in Figures 1a and 1b. These analyses indicated relationships between elements that were not apparent by comparing internal sequences. The results suggest that Mu3 and Mu4 were originated from MuDR (Mu9 ) by events independent from the ones leading to generation of the other subfamilies. The high similarity found among Mu7 and Mu6 TIRs agrees with the placement of these elements in the same subfamily. Despite its high level of internal homology with MuDR, Mu5 has TIRs that are no more closely related to those of MuDR than are those of the other defective Mu elements. This raises the possibility that the Mu1/Mu2 subfamily (for instance) may have been derived relatively recently from Mu5, and not directly from MuDR. The isolation of Mu8 from the other elements suggests that this element could have been originated directly from the autonomous MuDR, or from a MuDR-like element. As previously noted by many investigators, the two ends of a single element are usually most similar, implying that they evolve in a concerted manner possibly involving conversion across a cruciform structure.
The isolation of MuDR from the other elements suggests that either the intermediate defective elements are not present in the currently-existing maize populations or that such elements have not yet been discovered. Comparison of the degree of dissimilarity between TIRs of MuDR and of its defective relatives to the degree of sequence variation for the silent (i.e., intron and third position) sequences of adh1 (Gaut and Clegg, PNAS 90:5095-5099, 1993) suggests that MuDR diverged from its known defective elements less than ten million years ago. Since homologues of Mu elements have been found in species that diverged from maize more than 107 years ago, this suggests either horizontal transmission of these elements, concerted evolution of unlinked Mu element ends, or the loss/extinction (or a lack of detection to date) of defective Mu elements that are less closely related to MuDR. However, the application of molecular clock analysis to a transposable element (which may evolve more rapidly, by self-mutagenesis, or less rapidly, via concerted evolution, than standard maize genes) may be relatively inaccurate. The high dissimilarity between the TIRs of MuDR and other Mu TIRs has been suggested by Benito and Walbot (Maydica, 1994) to be a consequence of selection for promoter activity in the MuDR TIRs. Alternatively or additionally, there may have been selection against promoter function in the non-autonomous elements.
Figure 1a. Single Linkage Cluster Analysis
Figure
1b. Plot of the two principal components contributing to differences
between the TIR sequences
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}