The Suppressor-mutator (Spm) transposable element is subject to methylation-dependent epigenetic regulation in transgenic tobacco as it is in maize (Schläppi et al., Genetics 133:1009, 1993). The Spm promoter alone is rapidly methylated in tobacco (Schläppi et al., Cell 77:427,1994), but only if it contains the G+C-rich first untranslated leader sequence designated the 'downstream control region' (DCR) (Banks et al., Genes Dev. 2:1364, 1988). TnpA, the element's most abundant gene product, transcriptionally activates the methylated, inactive Spm promoter, but represses the unmethylated promoter (Schläppi et al., Cell 77:427, 1994). TnpA-mediated reactivation correlates with promoter demethylation. The DNA-binding and protein- dimerization domains of TnpA alone (Trentmann et al., Mol. Gen. Genet. 238:201, 1993) are sufficient for repression of the unmethylated promoter. By contrast, the C-terminus of TnpA, but not its N-terminus, is required for transcription activation of the methylated promoter. Only full-length TnpA, however, effects transposition of Spm elements in the presence of TnpD, another Spm-encoded protein required for transposition (Schläppi et al., Cell 77:427, 1994; Frey et al, EMBO J. 9:4037, 1990; Masson et al., Plant Cell 3:73, 1991). These results suggested that the C-terminus of TnpA directly activates transcription from the methylated Spm promoter and that both the N-terminus and the C-terminus of TnpA interact with TnpD to produce a transposition-competent complex. We have investigated these questions using both GAL4-based and a novel Spm-promoter-based single- and double-hybrid systems in transient plant cell assays.
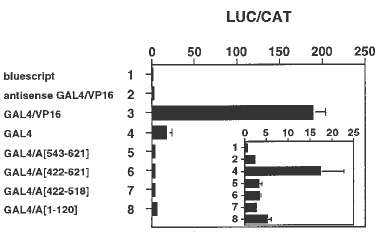
TnpA was tested for the presence of a transcription activation domain using TnpA-GAL4 DNA binding domain fusions (Giniger et al., Cell 40:767, 1985). The effector plasmid pPHI 1210, containing a translational fusion of the yeast GAL4 binding domain to the strong VP16 activation domain (from Herpes Simplex virus; Triezenberg et al., Genes Dev. 2:718, 1988), and the reporter plasmid DP 1446, containing a transcription fusion of GAL4 binding sites upstream of a minimal CaMV 35S promoter to a firefly luciferase gene (both kindly provided by Ben Bowen and Brad Roth, Pioneer Hi-Bred International, Inc.), were used to produce most fusion constructs. The VP16 activation domain in plasmid pPHI 1210 was replaced with different domains of TnpA to test for transcription activation potential (Unger et al., Plant Cell 5:831, 1993). As a background control, the VP16 activation domain was deleted from GAL4. Effector and reporter plasmids were co-introduced into tobacco suspension cells by microprojectile bombardment, and the results are shown in Figure 1. Compared to bluescript and GAL4/VP16 antisense control plasmids (lanes 1 and 2, respectively), the GAL4/VP16 plasmid activates the luciferase reporter gene about 100-fold (lane 3). Interestingly, the GAL4 binding domain by itself activates the reporter gene about 10-fold (lane 4). However, of the TnpA domains tested, neither the C-terminus by itself (amino acids [a.a.] 543 to 621), the C-terminus with the dimerization domain (a.a. 422 to 621), the dimerization domain by itself (a.a. 422 to 518), nor the N-terminus (a.a. 1 to 120) show any transcription activation (lanes 5, 6, 7, and 8, respectively). We conclude that TnpA lacks a conventional transcription activation domain.
TnpA binds to multiple copies of a 12-bp motif located in the element's promoter at the 5' end, as well as at the 3' end (Gierl et al., EMBO J. 7:2029, 1988). The last binding site in the promoter overlaps with the region at -30 upstream of the transcription start site (Pereira et al., EMBO J. 5:835, 1986). The Spm promoter is TATA-less and full promoter activity is associated with the presence of all TnpA binding sites contained within the 200-bp sequence termed the 'upstream control region' or UCR (Raina et al., Proc. Natl. Acad. Sci. USA 90:6355, 1993; Banks et al., Genes Dev. 2:1364, 1988). TnpA represses the unmethylated Spm promoter, possibly by competing with transcription factors for UCR binding (Schläppi et al., Cell 77:427, 1994). To test whether repression can be overcome by tethering a strong activation domain to the Spm promoter, plasmids containing translational fusions of VP16 to different TnpA domains were co-bombarded into tobacco suspension cells with luciferase reporter plasmids expressed from either the UCR alone or from the UCR and the G+C-rich DCR. The results are shown in Figure 2. Tethering of a strong activation domain to the Spm promoter indeed results in a strong activation of reporter gene expression, but only if the G+C-rich DCR is absent (compare lanes 4 and 6 with lanes 10 and 12, respectively). This result is consistent with the observation that the DCR confers enhancer-insensitivity on the Spm promoter (Raina et al., Proc. Natl. Acad. Sci. USA 90:6355, 1993). Consistent with the DNA-binding properties of TnpA, only fusions containing the DNA-binding domain (a.a. 120 to 422) activate the Spm promoter (compare lanes 2 and 8 with lanes 4, 6, 10, and 12). The construct including the TnpA dimerization domain (a.a. 1 to 543) is the strongest activator (lanes 6 and 12). TnpA constructs lacking the VP16 domain have no detectable background activity, in contrast to the GAL4 DNA binding domain, which itself stimulates LUC expression 10-fold (Figure 1, lane 1 compared to lane 4, and Figure 2, lanes 1, 3, 5, 7, 9, compared to lane 13). This is because TnpA is normally a transcriptional repressor of its own promoter and becomes an activator only when tethered to an activation domain. Thus, this new single-hybrid system consisting of the Spm promoter and the TnpA DNA binding domain can be used to measure transcription activation in transiently transfected tobacco cells with greater sensitivity than the GAL4 system provides.
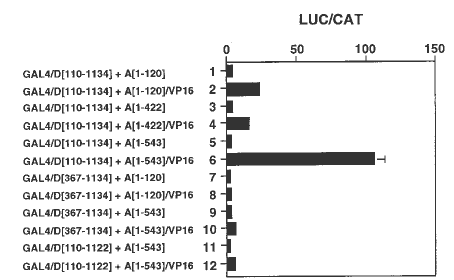
To determine which domains of TnpA interact with TnpD, we developed a plant double-hybrid system. TnpD domains lacking either the first 110 or 367 amino acids of the N-terminus (a.a. 110 to 1134 and 367 to 1134, respectively), or lacking the first 110 and the last 12 amino acids of the N- and C-terminus, respectively (a.a. 110 to 1122), were translationally fused to the GAL4 binding domain. The three TnpD-GAL4 constructs were individually co-bombarded into tobacco cells together with different TnpA/VP16 fusions and the luciferase reporter plasmid containing GAL4 binding sites. The results, shown in Figure 3, reveal that the VP16-containing N-terminus of TnpA (a.a. 1 to 120) can interact with a region between amino acid 110 and 367 of the N-terminus of TnpD, since deletion of this region prevents interaction of TnpA/VP16 and TnpD to activate the reporter gene (Figure 3, compare lanes 1 and 2 with lanes 7 and 8, respectively). The additional presence of the DNA-binding domain in TnpA (a.a. 1 to 422) had no effect on its interaction with TnpD (Figure 3, lanes 3 and 4). Inclusion of the protein dimerization domain in TnpA (a.a. 1 to 543) substantially increases the ability of the TnpA-VP16 fusion to activate luciferase expression (Figure 3, lanes 5 and 6; compare to TnpD lacking a.a. 110 to 367 in lanes 9 and 10). This observation implies that the TnpA-VP16 fusion protein dimer interacts more strongly with TnpD than the monomer. In addition, both the N-terminus of TnpD (a.a. 110 to 367), and the C-terminus are required for a productive interaction with TnpA. Deletion of only 12 amino acids from the C-terminus abolishes protein-protein interactions almost completely (Figure 3, lanes 11 and 12). By contrast, the first 110 a.a. can be deleted from the N-terminus of TnpD without loss of activity. Loss of TnpA-TnpD interaction is observed only with large N-terminal deletions. Thus both N- and C-termini of TnpD are required for TnpA-TnpD interactions, although only an intact C-terminus appears to be critical.
We have developed a new, highly sensitive, plant fusion protein system consisting of the Spm promoter and the Spm-encoded TnpA protein for detection of transcription activation domains in plant transient assays. We have also demonstrated the novel use of the yeast GAL4-Herpes Simplex VP16 system for detection of protein-protein interactions in a plant transient assay.
Figure 1. Analysis of putative TnpA transcription activation domains in transiently transfected tobacco cells. The strong transcription activation domain VP16 (from Herpes Simplex) in the effector plasmid pPHI 1210 was replaced with different TnpA domains from the N- or C-terminus to produce translational fusions with the GAL4 DNA-binding domain (from yeast). Each effector plasmid was co-bombarded individually into tobacco suspension cells together with the GAL4 binding site-containing luciferase (LUC) reporter plasmid DP 1446. To normalize for transfection efficiency, a chloramphenicol acetyltransferase (CAT) reporter plasmid was included, and relative promoter activity is expressed as the ratio of LUC to CAT activity. The LUC reporter plasmid is in each lane. The lanes are: Bluescript, pKS+ plasmid control (lane 1); antisense GAL4/VP16, antisense control (lane 2); GAL4/VP16, positive control (lane 3); GAL4, background control (lane 4); GAL4/A[543-621], a.a. 543 to 621of the C-terminus of TnpA fused to the GAL4 binding domain (lane 5); GAL4/A[422 -621], a.a. 422 to 621, the protein dimerization domain and the C-terminus of TnpA fused to the GAL4 binding domain (lane 6); GAL4/A[422 -518], a.a. 422 to 518, the protein dimerization domain of TnpA fused to the GAL4 binding domain (lane 7); GAL4/A[1-120], a.a. 1 to 120 of the N-terminus of TnpA fused to the GAL4 binding domain (lane 8).
Figure 2. Novel single-hybrid system based on the Spm promoter and the TnpA DNA-binding + dimerization domains for detecting transcription activation domains in tobacco cells. The effector plasmids have translational fusions of different TnpA domains to the VP16 activation domain. The reporter plasmids have LUC expressed from either the UCR (lanes 1 to 6) or from the UCR+DCR (lanes 7 to 13). In each lane, the CAT internal control is included, and promoter activity is expressed as the ratio of LUC to CAT activity. The lanes are: A[1-120], a.a. 1 to 120 of the N-terminus of TnpA (lanes 1 and 7); A[1-120]/Vp16, A[1-120] fused to VP16 (lanes 2 and 8); A[1-422], a.a. 1 to 422 of TnpA including the DNA-binding domain (lanes 3 and 9); A[1-422]/VP16, A[1-422] fused to VP16 (lanes 4 and 10); A[1-543], a.a. 1 to 543 of TnpA including protein dimerization domain (lanes 5 and 11); A[1-543]/VP16, A[1-543] fused to VP16 (lanes 6 and 12); bluescript, pKS+ plasmid control (lane 13).
Figure
3. Double-hybrid system based on GAL4/VP16 for analysis of TnpA and
TnpD interactions in tobacco cells. All cells received the LUC reporter
plasmid DP1446 (with GAL4 binding sites), and the CAT internal control
plasmid. Promoter activity is expressed as the ratio of LUC to CAT activity.
The lanes are: GAL4/D[110-1134], GAL4 binding domain fused to a.a. 110
to 1134 of TnpD (lanes 1 to 6); GAL4/D[367-1134], GAL4 binding domain fused
to a.a. 367 to 1134 of TnpD (lanes 7 to 10); GAL4/D[110-1122}, GAL4 binding
domain fused to a.a. 110 to 1122 of TnpD (lanes 11 and 12); A[1-120], a.a.
1 to 120 of the N-terminus of TnpA (lanes 1 and 7); A[1-120]/Vp16, A[1-120]
fused to VP16 (lanes 2 and 8); A[1-422], a.a. 1 to 422 of TnpA including
the DNA-binding domain (lane 3); A[1-422]/VP16, A[1-422] fused to VP16
(lane 4); A[1-543], a.a. 1 to 543 of TnpA including protein dimerization
domain (lanes 5, 9, and 11); A[1-543]/VP16, A[1-543] fused to VP16 (lanes
6, 10, and 12).
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}