Phytochemical mechanisms of resistance to ear rots
--D. J. Bergvinson and L.M. Reid
Recent studies have shown that both the kernel and silk possess phytochemical resistance mechanisms to Fusarium graminearum. Kernel resistance appears to consist of both physical resistance in the form of cell wall bound phenolic acids, and toxins in the form of soluble secondary metabolites. Silk resistance was initially thought to involve flavonoid biosynthesis in response to infection (Reid et al., Can. J. Bot. 70:1697-1702, 1992) but we have now identified resistant inbreds and hybrids with a contiguous wax layer covering the silk that is thicker than that found in susceptible genotypes.
During the past year we have been able to verify our field observations from last year's field season. Last summer we used scanning and transmission electron microscopy to see the morphological differences in the silk between resistant (CO272) and susceptible (CO266, CO265) lines. Compared to susceptible lines, CO272 had a visibly thicker wax coating on the silk as well as showing little or no checking in the silk.
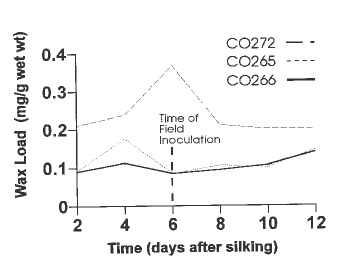
During the winter we extracted the silk wax to both quantify and characterize the wax load of CO272 and susceptible lines in a time-study experiment. We found CO272 to have four major wax constituents that ranged from 2 to 5 times the levels found in susceptible lines. In addition, we found the wax levels in CO272 to increase from the day of silk emergence to 8 days post-emergence and then drop off, while susceptible lines tended to peak at 4 days (Fig. 1). This trend is interesting from the field screening perspective, as the best spread in disease severity ratings is achieved when the silk channel inoculation is applied between 6 and 8 days post-silking (Reid et al. 1992).
Not only does wax load change over time but it also changes along the profile of the silk. Most genotypes tend to have the greatest wax load at the point where the silk emerges from the husk. In the case of CO272 the wax load is high along the entire length of the silk. We have also screened 66 Ontario commercial hybrids for wax load and compared this to field ratings. The most resistant hybrid (Pride K127) had the highest wax load of all the hybrids screened. However, unlike CO272, the commercial hybrid only had high levels of wax at the husk collar and the rest of the silk had levels comparable to more susceptible hybrids. This study to date has shown that wax load is at least a component, if not the main component, in silk resistance and that timings of sampling and positioning along the silk channel are important considerations when studying this resistance mechanism. We have now identified the major wax constituents to be simple hydrocarbons (C25-C31).
Last summer we conducted a time study for kernel resistance using our resistant Agriculture Canada inbred CO325 and a susceptible inbred CO266. We observed that kernel inoculation with F. graminearum resulted in the progressive spread to approximately 8 kernels (4 were wounded initially) in 5 days for both lines. However, by 9 days the spread had stopped for CO325 while the infection continued to spread in CO266 until the cob was completely infected by 28 days post-inoculation. We are presently doing phytochemical analysis on the kernels which were collected at 1, 3, 5, 7, 9, 11, 14, 21, 28, 35 days post-inoculation. Kernels have been fractionated into two fractions: pericarp/aleurone and germ/endosperm. Differences in soluble and cell-wall-bound phytochemicals over time and between lines will hopefully provide further insights into kernel resistance mechanisms.
During the winter we have developed a liquid bioassay procedure to test pure compounds that are found in corn for their effects on spore germination and hyphal growth of F. graminearum, F. moniliforme and F. subglutinans. The bioassay consists of 2 mL of Bilay's medium (1.5% sucrose) being added to 5 mL test tubes along with 10,000 spores and the test compound. After four days of periodic shaking, the contents of the tube are homogenized and the optical density recorded. Using this procedure we have found phenolic acids such as ferulic and p-coumaric acid to have an LC50 of approximately 200 ppm, while the most toxic compounds included salicylic and cinnamic acid. The phenolic amines did not demonstrate any toxicity using this bioassay system.
In summary, we have made progress in identifying waxes as a component in the silk resistance mechanism. Further work on how this mechanism is influenced by the environment is currently in progress. Kernel resistance appears to be most important two weeks after inoculation, and our efforts this season have focused on intensive sampling at this stage in ear development in hopes of seeing more dramatic differences between resistant and susceptible genotypes. We also have a bioassay system in place for testing compounds found in corn for their effects on spore germination and hyphal growth of the major Fusarium spp. from Ontario.
Figure
1. Time profile of silk wax load on resistant (CO272) and susceptible
(CO265 and CO266) corn lines.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}