The r-X1 deficiency is an X-ray induced submicroscopic deficiency that includes the r locus on chromosome 10. The deficiency is not transmitted through male gametes; however, it is transmitted with high efficiency through female gametes. If R/r-X1 plants (which are heterozygous for the deficiency) are testcrossed as female parents, about 55-60% of the kernels are colored (R/r) and 40-45% are colorless (r/r-X1) deficiency-bearing kernels. Plants germinated from the R/r kernels from the above cross are invariably diploid while those germinated from r/r-X1 kernels include 10-18% monosomics and 10-18% trisomics. The remaining plants are mostly diploids; however, multiply aneuploid plants and plants with deficiencies are also produced (reviewed in Weber, Maize Handbook ed. M. Freeling and V. Walbot, pp. 350-358, 1994). Thus, aneuploids are only produced from embryo sacs that contain the r-X1 deficiency. Clearly, a factor is present on chromosome 10 that is deleted in the r-X1 deficiency that is necessary for normal chromosomal disjunction during one or more of the embryo sac divisions. In the current study, we wanted to examine the effect of the r-X1 deficiency in the hemizygous condition in endosperm and embryo cells to determine if additional genetic factors could be assigned to the r-X1 deficiency.
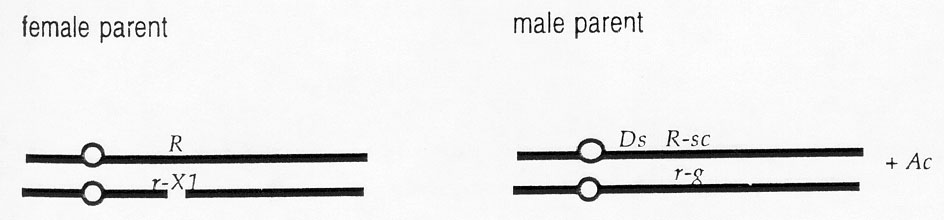
Production of endosperm sectors that are hemizygous for the r-X1 deficiency. Jerry Neuffer, University of Missouri, generously provided us with a stock that had a chromosome-breaking Ds that was proximal to the R locus (between the R locus and the centromere) on chromosome 10. Neuffer selfed an Ac-containing r-g/R-sc Ds4 plant and selected mottled kernels which were sent to us. These mottled kernels were produced when the Ds breaks the R-sc-bearing chromosome proximal to the R locus, the other homolog contains r-g, and Ac is present. In other words, the mottled kernels should have the same genotype as the original selfed Ac-containing r-g/R-sc Ds4 plant. The number of Ac elements that were present is not known. We crossed these as male parents on R/r-X1 plants in the inbred W22. This cross is diagrammed below (the r-X1 deficiency is shown as a gap in the chromosome):
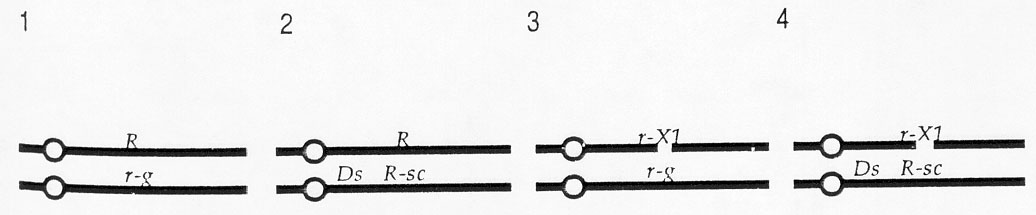
Kernels with the following four types of endosperms are expected (some have an Ac) (for simplicity, only one of the two identical maternally contributed chromosomes is shown):
Kernel types 1 and 2 are colored because they have a dominant allele of the R locus and kernel type 3 is colorless because it has one recessive r-g allele and a deficiency that includes the r locus. In kernel type 4, the Ds causes breaks in 10L proximal to the R locus if an Ac is present. Thus, the endosperms of these kernels should contain cells with the genetic constitution indicated above and some cells that are hemizygous for the r-X1 deficiency as diagrammed here:
Several different ears produced by the above cross each contained kernels with three different endosperm phenotypes. Type I kernels had purple normal endosperm, type II kernels had colorless normal endosperm, and type III kernels had purple etched endosperm. Type III kernels were usually somewhat smaller than the other two kernel types. We conclude that type I kernels had genotypes 1, 2, or 4 with no Ac, type II kernels had genotype 3, and type III kernels had genotype 4 with an Ac. Thus, it appears that kernel type III had a mixture of viable and inviable (or at least very slow-growing) cells to produce the etched phenotype, and we conclude that cells hemizygous for the r-X1 deficiency are inviable, and that a gene necessary for survival of endosperm cells has been lost in the segment that is deleted in the r-X1 deficiency.
Production of leaf sectors that are hemizygous for the r-X1 deficiency. We planted type I (purple, non-etched) and type III (purple, etched) kernels in our 1994 summer nursery. Leaves of plants grown from type I kernels were completely normal (no sectors); however, leaves of plants grown from type III kernels had numerous small sectors on them that were lighter green in color. Cells in the lighter green sectors appeared to be normal in size because the leaf surface was not altered in areas where a sector was present. No necrotic or albino areas were detected on plants grown from type III kernels. From these observations, we conclude that a locus is deleted in the r-X1 deficiency that is required for normal chlorophyll pigmentation in leaf cells; however, no locus required for leaf cell viability is deleted in the r-X1 deficiency. We had hoped to obtain large sectors on leaves of these plants that might be used for subtractive hybridization directed towards the cloning of genetic loci in the r-X1 deficiency; however, all of the sectors observed to date were extremely narrow (one or a few cells wide).
We conclude from this study that a locus required for endosperm viability and a locus required for normal chlorophyll pigmentation in leaves are deleted in the r-X1 deficiency in addition to the locus that is required for normal chromosome disjunction. We cannot be certain that the leaf and endosperm effects are caused by two different loci; however, this would seem likely.
Return to the MNL 69 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
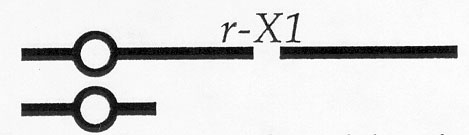{kind=link}