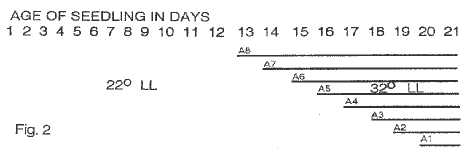
Under paramutagenic conditions for the R locus I reported in MNL, 1993, that it was possible to condition near colorless expression of the paramutant R gene in testcrosses of plants which as seedlings were held in constant light at 32 C for their first two weeks of development. Seedlings maintained in 22 C for three weeks showed significantly more pigmentation in their testcrosses at maturity. It was possible to bring about the near colorless level of paramutant R expression if seedlings which were started for two weeks in 22 C under constant light are shifted to 32 C and constant light in the third week. Figure 1 shows the testcross phenotypes of paramutated R-gene expression from R/R-lst plants which as seedlings received decreasing numbers of 32 C-LL cycles during the third week of seedling development. On all testcross ears, 50% of the kernels show different degrees of paramutant R expressions. Testcrosses labelled A8 through A1 represented plants which as seedlings received successively fewer days of the 32 C treatment. As seen in the lower half of Figure 1, with fewer than five days of 32 C and LL (A4-Al), a greater degree of variation in R-gene expression is encountered from ear to ear. Nevertheless, among all the treatments, A4 through A1, testcross ears are found whose R-gene expressions are as light as those from the treatments A8 to A5. An interpretation that can account for this heterogeneity is that the seedlings are not all receptive to the signal at the same time because of different stages of development. It could be inferred that a minimum of five days is necessary to have all plants respond to the applied temperature conditions. A corollary to this line of reasoning is that the change-over to the highly repressed state could be accomplished within a day or less if all seedlings were developmentally synchronized. Figure 2 shows, schematically, the treatment schedule for the first 21 days of seedling growth. Plants were kept in constant light for 21 days; beginning on day 13 the length of the line represents the number of days different seedling groups received 32 C-LL conditions. The only change throughout this three-week period was in the number of days seedlings were held in 32 C-LL conditions.
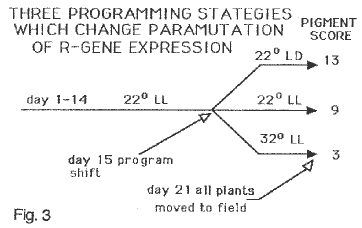
Figure 3 summarizes the programming strategies which can achieve different levels of paramutation as represented by the pigment scores to the right. Scoring was done as described in MNL, 1993. Unparamutated R genes score 18-20. LD (12 hr. light:12hr. dark) conditions administered the third week produced testcrosses with a pooled mean of 13; LL conditions for this same period produced plants at maturity whose testcrosses scored 9. Maximum reduction in pigment was achieved when seedlings were given 32 C for the third week of seedling treatment. For the first two weeks seedlings were held in constant light and 22 C. At the end of 21 days seedlings were moved to field conditions to complete their life cycle.
The significance of paramutation first reported by Brink in 1956 was that in a single generation a heritable change could be made in the expression of a Mendelian gene. All the R -gene expressions from the heterozygote with R-st were changed. A system which provided such a high frequency of responsiveness was an excellent place to see if the 20th Century dogma "the program does not learn from experience" (Jacob) could be challenged. The evidence outlined above and in previous issues of MNL shows that the genetic program learns from its genetic as well as its environmental experience. And, what is more important for a "good learner", the R gene has a "good memory" which can be incremented from generation to generation. Since the R gene controls a transcriptional activator and is responsive to both light and temperature, this memory capability should have important evolutionary significance for meeting the adaptive challenges demanded by the environment. Information processing, programmed for a specific stage of development, can now be thought of as a part of the regulatory apparatus of the paramutation system. The behavior of Spm elements as described by Fedoroff (Genetics 121:591-608 1989) shares some of the characteristics of the paramutation system described above.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}