During our studies on maize tissue culture, particular attention was focused on the development of embryogenic regenerable friable type II callus from elite maize genotypes and crosses of Lo inbred lines produced by the Section of Bergamo (Locatelli et al., MNL 66:17-18, 1992). The work was developed with the aim of evaluating the in vitro culturability of the Lo inbred lines, and establishing cultures for direct genetic transformation via electroporation in callus tissues as described by D'Halluin and coworkers (Plant Cell 4:1495-1505). In that paper, stable genetic transformation of maize was afforded by direct introduction of DNA into callus and immature zygotic embryos via electroporation. The advantage of that work, besides the importance of having established a new and easy tool for transformation, resides in the fact that DNA transfer can directly be applied to zygotic embryos after explant, or to type I primary calli, which can easily be induced in several maize genotypes (Hodges et al., Bio/Technol. 4:219-223, 1986).
On the other hand, we observed that the constant availability of immature embryos throughout the year needs a time consuming and sometimes problematic continuous breeding of donor plants in greenhouse or phytotron. Often, when greenhouse conditions can not be controlled perfectly, it is difficult to establish the optimal "physiological age" of the embryos to be explanted. Therefore genotypes that respond perfectly when evaluated from field conditions, do not respond as well when evaluated using embryos obtained from greenhouse grown plants. Furthermore, type I calli are easy to establish but difficult to maintain in culture for long periods, and their regenerative capability declines in a few months. A long term effort for the introduction of genes in maize would then be favored by a continuous supply of regenerable cultures, needing to be established only once a year. In this respect, we have chosen to direct our efforts to the establishment of optimal conditions for electroporation of selected embryogenic lines obtained from crosses of Lo inbreds with A188. Some selected lines behave as expected in that: i) they are friable, highly embryogenic and fast growing; ii) they are started yearly from immature embryos of the summer nursery; iii) they are easily maintained in propagation on N6P medium in the dark under standard routine conditions (as described in Lupotto and Lusardi, Maydica 33:163-177, 1988); iv) they are promptly regenerated after plating of calli in the light directly onto MS hormone-free medium, or after a 10 day culture on MS medium supplemented with 5 mg/l zeatin, in the light, and with subsequent transfer to hormone free conditions.
Besides maize we have established a variety of embryogenic callus cultures of cereal species: sorghum, bread wheat and durum wheat. Our objective is to develop a protocol of transformation in callus cultures via electroporation, amenable to utilization for gene delivery into the various species with minor modifications.
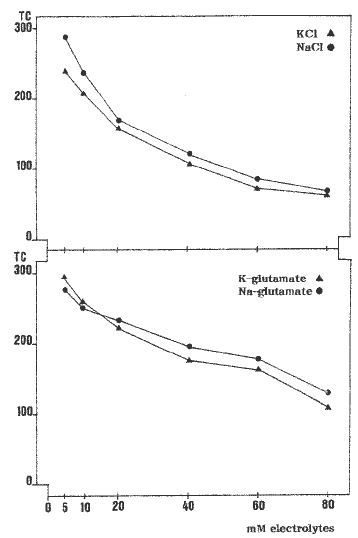
We have established a routine procedure for electroporation of maize type II calli by using a Bio-Rad Gene Pulser with capacitance extender, discharging one electrical pulse per sample with a field strength of 375-400 VÖcm-1 from a 960 uF capacitor. The callus used in each electroporation was about 80-150 mg fresh weight tissue chosen from the upper part of calli at the mid growth stage during subculture on N6P medium in the dark. In these conditions the tissue which is electroporated is mainly formed of globular shaped somatic embryos. Before electroporation, callus tissue is plasmolized for 2 hours at 24 C in 800 ul 0.4 M mannitol, 10 mM CaCl2ÖH2O, 10 mM MES, pH 7.2, with subsequent incubation 10 min on ice. After electroporation and a further incubation on ice for 10-15 minutes, callus pieces are transferred onto the surface of 0.6% agarose gelled N6P medium supplemented with 0.2 M mannitol, air dried for 30 minutes at the flow hood and incubated thereafter in the dark for subsequent growth. Various types of electrolytes can be utilized in the electroporation buffer such as KCl, NaCl, K-glutamate and Na- glutamate, in accordance with Songstad et al. (Plant Cell Tissue Organ Cult. 33:195-201, 1993). The duration of the electrical pulse, measured as time constant (TC) in msec, varies according to the electrolyte and its concentration, with stable inoculum size of callus tissue (Fig. 1). We could detect GUS expression in histochemically stained calli 48 h after electroporation in a range of values of TC, from about 70 msec, obtained with 80 mM KCl or NaCl, up to 300 msec, with 5-10 mM KCl or NaCl. Values of TC are slightly higher when Na-glutamate and K-glutamate are employed. Since our studies are focused on the stable introduction of genes into maize, a major requirement in the transformation procedure is represented by the ease and efficiency of regeneration of the electroporated tissues. We have noticed that when Na-glutamate and K-glutamate are utilized as electrolytes, independently from the concentrations, a strong decrease in the callus regenerative capability was observed. Furthermore, callus growth and somatic embryogenesis was also negatively influenced when calli were let grow on N6P medium. The highest regenerative efficiency was monitored when 10-20 mM NaCl was used for electroporation, and for this reason we currently utilize such conditions for stable transformation of maize. By using this procedure we have obtained stable transgenic maize callus lines containing marker genes, and work is in progress for plant regeneration and genetic analyses.
Figure
1. Electroporation of intact type II calli of maize. Electroporation
was in buffer containing two types of inorganic electrolytes (NaCl and
KCl) and two types of organic electrolytes (Na-glutamate and K-glutamate)
at various concentrations from 5 to 80 mM. TC is expressed in msec and
indicates time constant for a pulse discharged with 375 VÖcm-1 from
960 µF capacitor.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}