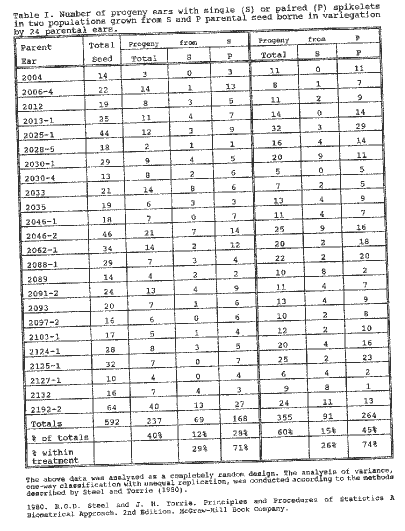
In last year's MNL, I reported that in certain of my connecting link stocks between teosinte and maize there was an unstable or variegated expression of the pd gene for single female spikelets. Being a non-molecular, old fashioned Mendelian geneticist, I have made a progeny test for the heritability of the differences between the single and paired spikelets on 24 ears variegated for these conditions. Every precaution was taken not to influence the data with bias. Several people, who were not directly interested in the results, were involved in the sorting of seed to plant, harvesting and scoring ears and analyzing the data as acknowledged. The results reveal a significant difference between the two populations, the one grown from single spikelet kernels and the other from paired spikelet kernels. The data flow sheet is in Table 1. The analysis of variance showing highly significant heritability of paired spikelets is in Table 2 and for single spikelets in Table 3.
Table 1. Number of progeny ears with single (S) or paired (P) spikelets in two populations grown from S and P parental seed borne in variegation by 24 parental ears.
Table 2. Number of paired spikelet ears from single (S) spikelet seed
and paired spikelet ears from paired (P) spikelet seed borne in variegated
ears.
|
|
ri | Yij | meanYi | Y^2ij | Y^2i/ri | (Yij-meanYi)^2 |
| S | 237 | 168 | 0.71 | 1846 | 119.1 | 1619.54 |
| P | 355 | 264 | 0.74 | 3970 | 196.3 | 3592.42 |
| sum | 592 | 432 | 5816 | 315.4 | 5211.96 |
Analysis of variance for data summarized above
| Source of variation | df | SS | MS | F |
| Among treatments | 1 | 288.84 | 288.84 | 32.71** |
| Within treatments | 590 | 5211.96 | 8.83 | |
| Total | 591 | 5500.8 |
The analysis of variance given above is significant; tabulated F(0.005)=7.88 for 1 and 590 degrees of freedom. Therefore sufficient evidence exists to suggest that the mean number of paired spikelet types produced from single (0.71) and paired (0.74) spikelet type parent plants are different for reasons other than chance.
Because both the single and paired conditions on these variegated ears are inherited and because a pair of spikelets produces twice as many kernels as the single ones, the population would of its own accord shift to the paired condition without human help.
Table 3. Number of single spikelet ears from single (S) spikelet seed
and single spikelet ears from paired (P) spikelet seed borne in variegated
ears.
| Trt | ri | Yij | meanYi | Y^2ij | Y^2i/ri | (Yij-meanYi)^2 |
| S | 237 | 69 | 0.29 | 407 | 20.09 | 369 |
| P | 355 | 91 | 0.26 | 561 | 23.33 | 515.36 |
| sum | 592 | 160 | 968 | 43.42 | 884.36 |
Analysis of variance for data summarized above
| Source of variation | df | SS | MS | F |
| Among treatments | 1 | 40.4 | 40.4 | 26.93** |
| Within treatments | 590 | 884.36 | 1.5 | |
| Total | 591 | 924.76 |
The analysis of variance given above is significant; tabulated F(0.005)=7.88 for 1 and 590 degrees of freedom. Therefore sufficient evidence exists to suggest that the mean number of single spikelet types produced from single (0.29) and paired (0.26) spikelet type parent plants are different for reasons other than chance.
The data clearly demonstrate at least an inherited component, possibly of a transposon nature, that is regulating pd-Pd expression and is passed on to the next generation. The random distribution of paired and single spikelets is mostly in the central region of the ear with more paired ones near the base and more single ones high on the ear. If we assume that it takes more energy (glucose) to differentiate a pair of spikelet primordia than just a single one, this pattern would fit. But even so, this does not exclude transposon regulation. It is generally agreed that the internal environment of the host controls transposon movement during morphogenesis. Perhaps in this case it is the rhythms of distribution of photosynthate that stem from day-night cycles, just as plastochrons of phytomers do, that influence transposon movement which then regulates gene activity. In any case, whether or not energy level is involved, the pd-Pd states of expression in a variegated arrangement are inherited. I previously reported a similar situation regarding floral and vegetative multiranking (mr-Mr). In this case there was increased transmission of full vegetative multiranking through pollen from the central spike of the tassel in comparison with pollen from the two-ranked lateral branches (MNL 64:120, 1990).
Acknowledgments: Dr. Ann E. Kennedy for carefully separating kernels borne in paired spikelets, even when only rudiments of the second spikelet were apparent, from those kernels borne only one per cupule, and putting them in planting envelopes. Dr. Neelima Sinha for planting this single spikelet seed and paired spikelet seed from 24 divided ear-to-row families. Mr. Miguel Sosa and Mr. Oscar Hernandez for harvesting the uppermost ear on the main stalk from each plant and labeling these ears for classification. Bill Ebener of Mesa Inc. for crucial help with the statistical analysis.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}