Plants use sunlight for photosynthesis, and are unavoidably exposed to the ultraviolet radiation that is also present in sunlight. Because plants lack the behavioral mechanisms that mobile animals use to respond to excess radiation, they may have evolved novel and effective mechanisms to protect essential functions from radiation damage. Plants have two basic mechanisms for coping with UV damage--shielding and repair.
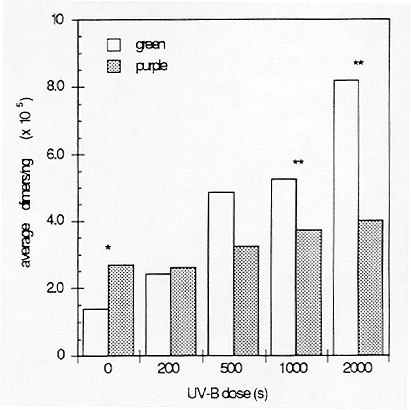
Between 95% and 99% of incoming UV radiation does not penetrate through the epidermal layer. Most of this UV shielding is due to absorption by epidermal flavonoid compounds (Robberecht and Caldwell, 1978, Oecologia 32:277). We show that anthocyanins synthesized under the control of the regulatory genes B and Pl can protect maize leaf DNA from damage induced by UV radiation in vivo (Fig. 1).
Thus the interior of the plant is protected from the induction of UV radiation damage by epidermal anthocyanins. Shielding, however, does not provide complete protection from UV damage, and is not sufficient to explain differences in UV tolerance between different plants (Sullivan et al., Amer J Bot 79:737, 1992).
We have therefore measured the repair of UV-induced DNA damage in maize seedlings. We have measured both photoreactivation repair (in which the enzyme photolyase uses light to remove UV-induced DNA damage) and excision repair (which occurs in the light and in the dark).
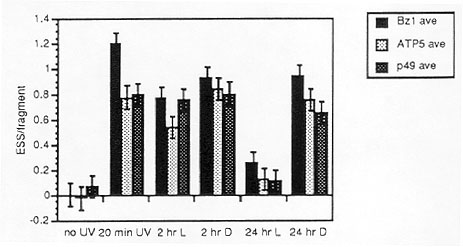
We demonstrate (Fig. 2) that there is photoreactivation-type repair (reduced damage after 24 hrs. of recovery in the light) of UV-induced DNA damage in nuclear, mitochondrial and chloroplast genes in maize seedlings. This suggests that all three compartments contain photolyase. Developmental and molecular characterization of maize photolyase(s) is proceeding.
Figure 1. DNA damage levels in green (b pl r-g) and purple (B Pl) plants after UV-B irradiation. Sheath tissue was irradiated, DNA extracted and amount of cyclobutane pyrimidine dimer damage measured using our antibody assay (Stapleton and Walbot, Plant Mol. Biol. Rep. 11:230, 1993).
Figure
2. Repair of UV-induced DNA damage in maize seedlings. Seedlings (bz2/bz2)
were grown for 1 week, irradiated with 6 germicidal UV-C bulbs for 20 min.,
and allowed to recover for 2 or 24 hrs. in light or dark. Damage was measured
using a technique originally developed for determination of repair rates
in mammalian tissue cultures (Bohr and .i.Okumoto, in DNA Repair,
a Laboratory Manual, E. C. Friedberg and P. C. Hanawalt, eds., Marcel Dekker,
New York;, 1988). We determined damage levels by determination of the number
of sites in each HpaI fragment sensitive to T4 endonuclease V (ESS/fragment)
that hybridized to a nuclear (Bz1), chloroplast (ATP5) or mitochondrial
(p49) probe.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}