QTL (quantitative trait locus) mapping identified the region between umc18 and umc60 on chromosome arm 3L as affecting several aspects of inflorescence structure that distinguish maize and teosinte (Doebley and Stec, Genetics 134:559-570, 1993). These include (1) the extent to which inflorescences terminating the primary lateral branches are male or female, (2) inflorescence phyllotaxy, (3) the length of the internodes in the primary lateral branches, (4) the frequency of paired versus single spikelets in the female inflorescence, and (5) the number of cupules (internodes) within the inflorescence. One possibility is that all of these differences represent the pleiotropic effects of a single locus located within this segment. Previously, I proposed that this locus might be terminal ear1 (te1) (Doebley, Cell. Biochem., Supl. 17B, p. 5,1993). In this note, I explain this hypothesis and present new evidence consistent with it.
A model for terminal ear1 in maize evolution. Terminal ear1 plays a role in internode elongation in the vegetative culm. Mutant te1-ref plants have shorter internodes in the vegetative culm of the plant and often some female spikelets in the tassel. The internodes just below the tassel are the most severely affected relative to wild type. The model presented here presumes that, in teosinte, te1 plays a role in internode elongation in both the vegetative culm and in the inflorescences. Then, during the evolution of maize, te1 was altered such that the maize allele (Te1-Maize) produces shorter internodes in the inflorescences and the primary lateral branch (or ear shank) than does the teosinte (progenitor) allele (Te1-Teosinte). One mechanism by which this could be accomplished is if Te1-Maize has a lower level of expression in the inflorescence than does Te1-Teosinte. Shorter internodes in the ear and ear shank would have pleiotropic effects on phyllotaxy, inflorescence sex, and single versus paired spikelets. Also, if the internodes are shorter in the ear, then it may be possible to produce a larger number of internodes. Under this model, both the maize and teosinte alleles of te1 would produce functional products. Finally, since te1-ref represents a loss of function mutation, it may also cause shorter internodes in the ear shank and the inflorescence. These effects may be difficult to detect because internodes in the inflorescences and ear shank are already very short in most types of modern maize.
New evidence. Two new pieces of information are consistent with the proposal that te1 is our QTL. First, I scored internode length in the ear as a quantitative trait in a maize-teosinte F2 population. The ears of the F2 plants showed a range of variation from those in which all internodes were relatively long and of equal length (like teosinte) to others with alternating short and long internodes (Fig. 1). The later condition is both more maize-like and reminiscent of the effect that te1 can have on the vegetative culm. Interval mapping revealed seven QTLs affecting internode length in the ear (Table 1). The QTL with the largest effect accounts for 47% of the variation (a very large effect) and maps to chromosome arm 3L where te1 is located. Substitution of two maize alleles at this QTL for two teosinte alleles reduces the average length of the internodes in the ear from 4.5 to 3.0 millimeters.
Table 1. Location and effects of quantitative trait loci affecting internode
length in the ear based on an analysis of 290 F2 plants derived from a
maize (race Reventador) by teosinte (ssp. parviglumis) cross. This
population previously described by Doebley and Stec (Genetics 134:559-570,
1993).
|
|
Chromosome | LOD-score | R-squared | Candidate |
| umc157 | 1S | 6.06 | 12.1 | |
| umc107 | 1L | 2.63 | 4.4 | tb1 |
| umc5a | 2L | 2.42 | 4.1 | |
| umc60 | 3L | 19.19 | 47.0 | te1 |
| umc42 | 4S | 10.68 | 26.2 | tga1 |
| umc27 | 5S | 2.47 | 4.6 | |
| umc110a | 7L | 5.71 | 12.0 |
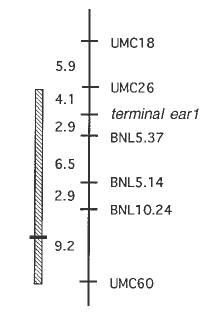
A second piece of supporting evidence was possible because te1 has recently been molecularly cloned (Bruce Veit, pers. comm.). Using a clone of te1 (generously provided by B. Veit and S. Hake), I was able to map te1 relative to our QTL. te1 is located in the interval between umc18 and umc60 where our QTL for inflorescence internode length maps (Fig. 2). These new data make te1 a strong candidate for our QTL. The recent cloning of te1 makes it possible to test definitively both the hypothesis that our QTL is te1 as well as the specific predictions of the model outlined above.
Finally, it is interesting that two of the other QTLs, affecting internode length in the ear, map near other genes involved in the evolution of maize (Table 1). There is a QTL of small effect on 1L near tb1. Our analysis of tb1 indicates that it primarily affects internode length in the ear shank but that it also has some effect on internode length in the ear itself (Doebley and Stec, unpublished). A second QTL of large effect maps near tga1 on 4S. Dorweiler et al. (Science 262:233-235, 1993) have reported that one of the effects of tga1 is on the length of the internodes within the ear.
Figure
1. Ears from a maize-teosinte F2 population showing variation in the
length of internodes within the ear: teosinte-like ear with internodes
that are long and of relatively equal lengths so that the cupulate fruitcases
appear to be stacked one on top of the other (left), and more maize-like
ear with alternating short and long internodes so that the cupulate fruitcases
appear side-by-side (right).
Figure
2. Map of a portion of chromosome arm 3L. Hatched rectangle on the
left represents the 2-LOD support interval for the position of the QTL
affecting internode length in the ear.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}