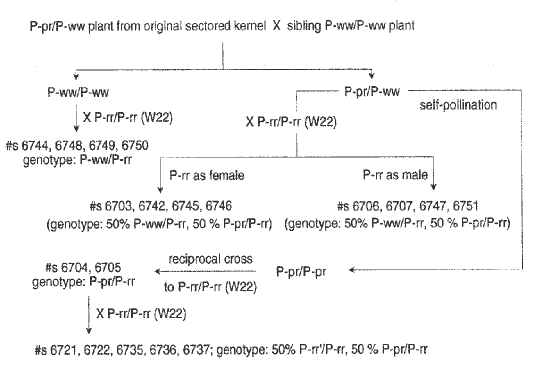
The preceding note demonstrates that P-pr reduces the pigmentation caused by P-rr, indicating that the alleles interact. Here we present evidence for heritability of interaction, determined as modification of the phenotype of P-rr' (in analogy with the convention for paramutation at R and B loci) transmitted after interaction with P-pr. The genetic scheme used is shown in Figure 1. Test plants of the genotype P-pr/P-rr, and control plants of genotype either P-ww/P-rr or P-rr'/P-rr were testcrossed to P-ww. The resulting kernels were planted in blocks that yielded between 90 and 160 plants, and phenotypes were determined in the field on open-pollinated ears. Phenotypes were classified as white (colorless, expected from P-ww/P-ww), red (solid red color on all kernels, expected from P-rr/P-ww), patterned (variegated pigmentation on all kernels, expected from P-pr/P-ww) and a novel phenotype designated variegated red, which was largely red, but resembled the P-pr phenotype on some or all kernels of an ear. The distinction between variegated red and patterned was somewhat subjective, since it was based on the criterion of which color, red or white, was more predominant on the ear. However, this does not affect the conclusions, as shown below.
Table 1. Segregation in testcrosses to P-ww/P-ww (4Co63).
| Control populations of genotype P-ww/P-rr |
| Family # (see Fig. 1) | # plants crossed | Red ears | White ears | Variegated ears | Chi-square |
| 6744 | 4 | 301 | 308 | 0 | 0.08 |
| 6748 | 5 | 333 | 333 | 1 | 0 |
| 6749 | 7 | 304 | 362 | 0 | 5.05 |
| 6750 | 3 | 189 | 197 | 0 | 0.17 |
| 6703 | 6 | 294 | 306 | 0 | 0.24 |
| 6742 | 2 | 147 | 113 | 0 | 4.45 |
| 6706 | 15 | 1004 | 1079 | 0 | 2.7 |
| 6707 | 4 | 322 | 310 | 0 | 0.23 |
| Total | 46 | 2894 | 3008 | 1 | 2.2 |
| Test populations of genotype P-pr/P-rr |
| Family # (see Fig. 1) | # plants crossed | Red ears | Patterned ears | Variegated ears | Chi-square |
| 6735 | 10 | 602 | 602 | 41 | 1.35 |
| 6736 | 4 | 167 | 224 | 43 | 23 |
| 6737 | 6 | 50 | 526 | 155 | 544.7 |
| 6722 | 3 | 189 | 190 | 2 | 0.02 |
| 6721 | 2 | 121 | 110 | 20 | 0.32 |
| 6703 | 9 | 514 | 505 | 31 | 0.46 |
| 6704 | 32 | 1658 | 2098 | 185 | 99.1 |
| 6705 | 24 | 1174 | 1666 | 211 | 162 |
| 6706 | 10 | 555 | 664 | 66 | 23.8 |
| 6707 | 6 | 394 | 472 | 51 | 18.1 |
| 6742 | 1 | 63 | 65 | 1 | 0.07 |
| 6745 | 5 | 284 | 379 | 44 | 27.3 |
| 6746 | 4 | 331 | 333 | 8 | 0.15 |
| 6747 | 2 | 145 | 160 | 9 | 1.83 |
| 6751 | 2 | 148 | 192 | 9 | 2.2 |
| Total | 120 | 6395 | 8186 | 876 | 460.7 |
Segregation data for the families described in Figure 1 are shown in Table 1. Testcrosses to P-ww of control populations of genotype P-ww/P-rr gave 5903 ears from 46 subfamilies, representing 8 progenitor families. Virtually all ears were red or white; the single variegated red ear may be due to pollen contamination. Testcrosses to P-ww of a second control population of genotype P-rr'/P-rr yielded 2937 ears from 22 subfamilies, representing families 6721, 6722, 6735, 6736 and 6737 (Fig. 1); these were also all solid red, with no variegated red ears (data not shown in the Table). In contrast, almost all of the 15 test families of genotype P-pr/P-rr gave a significant number of variegated red ears, in addition to patterned red and solid red ears. A few white or almost white ears were also seen, which, if confirmed for the presence of a new P-ww allele, may represent loss of function derivatives of P-pr or P-rr.
The presence of the variegated red class in testcrosses of P-pr/P-rr may result either from an increase in pigmentation capacity of P-pr, or from a decrease in pigmentation capacity of P-rr'. These alternatives were tested by analysis of segregation ratios. The first alternative predicts that the sum of variegated and patterned ears should equal the number of red ears, while the second predicts that they should exceed it. Chi-square analysis of the departure from 1:1 segregation for both control and test populations is shown in Table 1 (underlined values are significant at the 95% level). In the control population, only two families deviate significantly from a 1:1 ratio of red and white ears, and these show opposing biases. The total shows no significant deviation, indicating that the genetic background or unlinked factors do not lead to consistently biased segregation. In contrast, in the test population, 7/15 families deviate at the 95% confidence level from 1:1 segregation of the patterned + variegated class relative to the solid red class. All of these represent decreases in the solid red class, leading to a highly significant chi-square value of 461 for the total.
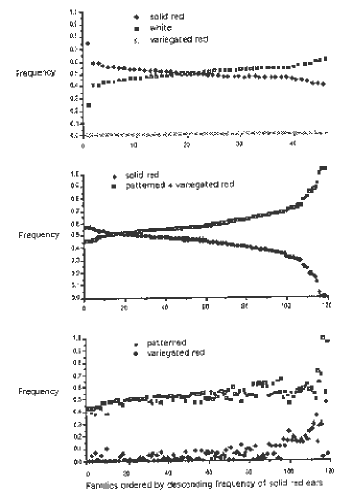
Figure 2 shows the same data for each subfamily, ordered by decreasing frequency of red ears. For the control population (upper panel), all values adhere fairly closely to the expected value of 0.5, and deviations cancel out as indicated by the fact that the two curves cross close to the middle. In contrast, for the test population, segregation values are distorted, with fewer solid red ears in most families, as indicated by the crossing of the two curves near the left end. Progeny from the ears represented on the right-hand side of the range are highly deficient in red ears, suggesting nearly complete loss of the normal pigmentation capacity of P-rr. These observations indicate a similarity in the interaction between these two alleles to paramutation at the R locus.
Figure
1. Genetic Scheme: the chart shows the lineage of the families whose
members were testcrossed to P-ww to obtain the data in Figure 2
and Table 1.
Figure
2. Segregation in Control and Test Populations: Segregation frequencies
from the 46 subfamilies of the control population (upper panel) and 120
subfamilies of the test population (middle and lower panels) were ordered
by decreasing frequency of red ears, and plotted.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}