Aspartate kinase activity extracted from seedlings of the ask1
mutant
--Ricardo A. Azevedo and Peter J. Lea
The growth of cell cultures and seedlings of many plants can be inhibited by lysine plus threonine due to feedback inhibition at one or more steps in the aspartic acid metabolic pathway (Green and Phillips, Crop Sci. 14:827, 1974). The selection of mutants showing resistance to the inhibition caused by lysine plus threonine has shown that less sensitivity to feedback regulation leads to an overproduction of soluble threonine (Hibberd and Green, PNAS 79:559, 1982). In barley, lysine plus threonine resistant mutants showed that mutant forms of aspartate kinase isoenzymes were less sensitive to lysine feedback inhibition (Arruda et al., Plant Physiol. 76:422, 1984).
In maize, mutants resistant to inhibition by lysine plus threonine were also obtained and the enzyme aspartate kinase was extracted from maize ears and cell culture and analysed (Dotson et al., Planta 182:546, 1990). The ask1 gene, which is the structural gene for aspartate kinase, was transferred to the Cat100-1 inbred line, mapped in the short arm of chromosome 7, and shown to be regulated by the opaque2 gene when soluble and total amino acid fractions were analysed (Azevedo et al., Plant Sci. 70:81, 1990). This work represents a continuation of investigations on the ask1 mutant gene.
The mutant ask1 gene was transferred from line A619 to Cat100-1 by backcrossing to near isogenic conversion. Maize seeds from ears containing separately Ask1/Ask1 and ask1/ask1 genotypes were planted in trays containing organic compost, incubated at 28 C in a 16/8 h light/dark period for 10 days, and water added at 48 h intervals. At the end of this period (seedlings normally showing 5 leaves) the leaves were harvested and the weight recorded. The leaf samples were immediately frozen with liquid nitrogen and used for aspartate kinase extraction.
For the identification of the presence of the ask1 gene in the homozygous dominant form, 40 seeds from each of 8 segregating ears were planted and grown to maturity in a greenhouse (Lancaster University Field Station) at 24 C. Anther spikelets from each plant were collected and stored at -80 C. Anthers were then used for extraction of soluble amino acids. One anther spikelet from each plant was homogenized with 30 µl of distilled-deionised water in 0.5 ml microfuge tubes using a glass rod. The homogenates were centrifuged in a microcentrifuge at 16000 rpm and the supernatant used for amino acid analysis. Aliquots of 5 µl were applied to thin-layer chromatography (TLC) on glass plates (20x20 cm) coated with 0.5 mm of a 2:5 (w/w) mixture of silica gel and cellulose. The chromatogram was developed in a solvent mixture containing butanol, acetone, ammonium hydroxide and water (5:5:2.5:1, by volume) for 90 min. After the separation of the amino acids, the plates were dried for 3 days at room temperature and sprayed with 0.2% (w/v) ninhydrin in acetone. Standard threonine (1%, w/v) solution in water was used to identify the amino acid spots.
The segregation of the ask1 gene was recorded and seeds from ears homozygous (Ask1/Ask1) for the gene were selected for the experiment.
Aspartate kinase was extracted and partially purified from 10 g of tissue for each genotype. Samples were ground with liquid nitrogen in a pestle and mortar with 5:1 (v/w) extraction buffer to 5% (w/v) polyvinylpyrrolidone. The extraction buffer contained 50 mM Tris/HCl ph 7.4 with 50 mM KCl, 2 mM lysine, 2 mM threonine, 1 mM DTT, 0.1 mM PMSF, 20% (v/v) ethanediol and the extract was filtered through several layers of gauze. After ammonium sulphate precipitation (35-60%), protein pellets were resuspended and desalted on G25 Sephadex columns equilibrated with Tris buffer (pH 7.4) containing 50 mM KCl, 0.1 mM lysine, 0.1 mM threonine and 10% (v/v) ethanediol. Aspartate kinase activity was measured by the hydroxamate assay method (Azevedo et al., Phytochemistry 31:3725, 1992).
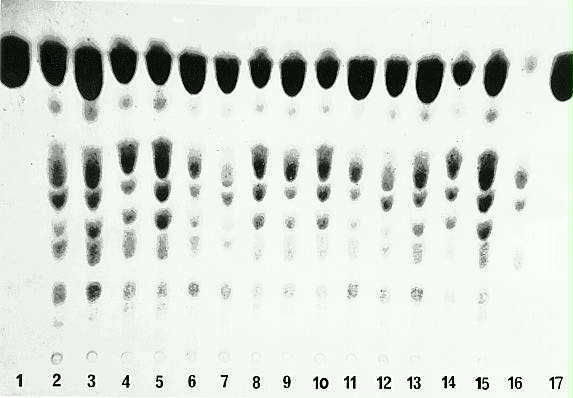
The identification of the ask1 gene had to be carried out since the seeds were segregating (Ask1/Ask1:Ask1/ask1:ask1/ask1) for the gene. A large number of seeds from each of the 8 ears were selected and planted, and grown to maturity, producing panicles from which anther spikelets were collected and tested. Some of the plants were self-pollinated or crossed. The extraction of soluble amino acids from anther spikelets and their identification and quantification by TLC has been shown to be a reliable indicator of the presence of the ask1 gene. This was also the case in these experiments, since very clear differences in the levels of threonine among the genotypes could be observed (Fig. 1). From 8 ears tested, only ear number 7 showed the gene in the homozygous dominant form (Ask1/Ask1) with all plants being threonine overproducers. The analysis of the other 7 ears showed that two of them, 3 and 4, were normal (ask1/ask1) whereas ears 1, 2, 5, 6 and 8 showed segregation for the gene (Table 1). This experiment was carried out with the objective of identifying the ask1 gene, therefore no quantitative measurements were carried out to separate Ask1/Ask1 from Ask1/ask1 since both genotypes overproduce threonine. Table 2 shows the results obtained when aspartate kinase activity was determined in wild type (ask1/ask1 ) and in the mutant (Ask1/Ask1) plants. Aspartate kinase was only slightly higher in the mutant than in the wild type plants. However, variation in the inhibition caused by lysine was observed. Lysine (5 mM) produced a 65% inhibition of aspartate kinase activity in the wild type, while the same concentration of lysine showed a 41% inhibition in the mutant. Threonine, on the other hand, did not show any variation, giving identical levels of inhibition of aspartate kinase activity between the two genotypes tested. This result confirmed the results reported by Dotson et al. (Planta 182:546, 1990), who analysed two mutations (one of them the ask1 gene) in which aspartate kinase activity was isolated and purified from immature maize ears. However, the extent of the lysine inhibition was different. This may be due to the fact that a different tissue was used, which might alter the effect of these amino acids on the enzyme activity. These results also confirmed that the ask1 gene is one of the structural genes for the lysine-sensitive aspartate kinase in maize. Although Dotson et al. (Planta 182:546, 1990) could not identify the threonine-sensitive form of aspartate kinase, these results strongly support the affirmative above, since the inhibition by threonine was not altered in the mutant. Thus other genes may be responsible for the threonine-sensitive aspartate kinase isoenzyme in maize.
The addition of lysine and threonine together showed a clear additive pattern of these two amino acids on the aspartate kinase activity. In the mutant, the addition of lysine plus threonine did not show a strong aspartate kinase inhibition as observed for the wild type due to the reduced sensitivity of the lysine-sensitive aspartate kinase. However, this reduction was smaller than the levels expected for an additive effect of the two amino acids as shown by wild type plants.
Additional experiments with anthers, seedlings and endosperms are being carried out to better characterize the ask1 gene in maize.
Table 1. Segregation of the ask1 gene in 8 progenies obtained
from self-pollination of Ask1/ask1. The segregation of the gene
was verified by the threonine levels of different genotypes in TLC plates.
|
|
|||||
|
|
|
|
|
|
|
| 1 | 40 | 7 | 5 | 2 | Ask1/ask1 |
| 2 | 40 | 4 | 3 | 1 | Ask1/ask1 |
| 3 | 40 | 11 | 0 | 11 | ask1/ask1 |
| 4 | 40 | 7 | 0 | 7 | ask1/ask1 |
| 5 | 40 | 9 | 6 | 3 | Ask1/ask1 |
| 6 | 40 | 5 | 4 | 1 | Ask1/ask1 |
| 7 | 40 | 12 | 12 | 0 | Ask1/Ask1 |
| 8 | 40 | 15 | 11 | 4 | Ask1/ask1 |
Table 2. Aspartate kinase activity (nKat/mg protein) extracted from
wild type and mutant seedlings for the ask1 gene.
|
|
||||
|
|
|
|||
|
|
|
|
|
|
| Control | 0.0141 | 0 | 0.0153 | 0 |
| Thr (2 mM) | 0.0104 | 26 | 0.0112 | 27 |
| Thr (5 mM) | 0.0097 | 31 | 0.0109 | 29 |
| Lys (2 mM) | 0.0062 | 56 | 0.0109 | 29 |
| Lys (5 mM) | 0.0049 | 65 | 0.0090 | 41 |
| Lys+Thr (2 mM) | 0.0028 | 80 | 0.0076 | 50 |
| Lys+Thr (5 mM) | 0.0015 | 89 | 0.0067 | 56 |
Figure
1. Thin layer chromatograms of amino acids extracted from anthers for
the identification of the ask1 gene, which leads to overproduction
of threonine. Lanes 1 and 17 correspond to the threonine standard; 2-15
correspond to different plants showing threonine accumulation and 16 corresponds
to the negative control (wild type).
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}