Selection of plants resistant to S-2-aminoethyl-L-cysteine
--Ricardo A. Azevedo and Paulo Arruda
Substantial progress has been made with genetic, biochemical and molecular techniques in the study of metabolism and development of plants. Plant tissue culture techniques have been widely used and have proved to be a useful tool for the induction, selection and study of mutants. They have also brought new insight into the role of enzymes involved in primary metabolism. However, for such an approach to be viable, protocols are needed for the establishment of callus, cell suspension and protoplast cultures, as well as plant regeneration. Furthermore, strategies for the in vitro selection of mutants are fundamental.
Mutants selected in vitro with an altered aspartate kinase (AK) have been obtained in maize, leading to an overproduction of threonine in different plant tissues, including endosperm (Hibberd and Green, PNAS 79:559, 1982; Azevedo et al., Plant Sci. 70:81, 1990; Diedrick et al., TAG 75:209, 1990). The lysine analog S-2-aminoethyl-L-cysteine (AEC) has been used to select mutants with an altered dihydrodipicolinate synthase (DHDPS), which is strongly inhibited by lysine and AEC. The relaxation of this regulatory step of the pathway could lead to an accumulation of soluble lysine in the maize seed.
Maize tissue culture was used to select plants resistant to AEC. Calli treated with NaN3 and selected in MO9-2 medium containing 0.25 mM AEC showed the formation of necrotic and non-necrotic sectors. Normally growing sectors, when subcultured continuously into the same medium, showed resistance to AEC and were then transferred to the regeneration medium. Although normal callus growth has been observed after several subcultures in the presence of AEC, a small number of regenerating plants did not survive the AEC inhibition of root growth in the regeneration medium. However, 173 plants were regenerated, from which 63 reached the field stage and 35 of these reached maturity with the production of a panicle and ears, which allowed self-pollination and crossings to the original Cat 100-1 inbred line.
The mutagenic NaN3 proved to be very efficient by the number of mutants observed. Meanwhile, the selection system still allowed a large number of escapes, since many mutants not related to AEC inhibition were obtained, as well as plants showing no alteration. This could be due to a gradient effect of AEC produced by a differential uptake of AEC by cells in the upper layers of the calli, where most of the normal growing cells were observed. Thus, successive subcultures of these normal sectors during the selection stage for a shorter period of time might reduce the number of undesirable mutants. Furthermore, the use of cell suspension and protoplast cultures could also show a better efficiency; however, even using cell suspension cultures, escapes are still a common feature, with at least 25% sensitive plants.
Anther spikelets from each regenerated plant were tested for soluble amino acid overproduction with special attention to lysine. Chromatograms showed a large variation in the concentration of different amino acids, including lysine. Plants AEC-5, AEC-6, AEC-11, AEC-8, AEC-14, AEC-23 and AEC-25 showed strong bands with the same Rf as that of lysine, while the control Cat 100-1, as well as other regenerated plants tested, did not show a band which corresponds to lysine.
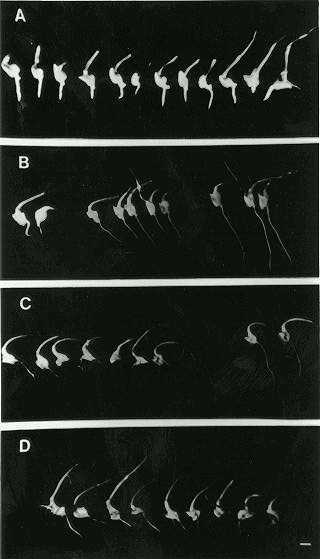
Progenies obtained from regenerated plants were tested for resistance to AEC by inoculating excised immature embryos in the presence of AEC (Fig. 1). Among all progenies tested, only two, AEC-1 and AEC-5, showed segregation for AEC resistance.
Genetic analysis showed that for the AEC-1 resistant mutant, the resistance was a dominant trait segregating according to Mendelian laws (Fig. 1B). For the AEC-5 resistant mutant, on the other hand, the resistance was a recessive trait (Fig. 1C). These results were further confirmed when F1 progeny were produced by crossing AEC-1 (in the recessive homozygous condition, aec-1/aec-1) and AEC-5 (in the dominant homozygous condition, Aec-5/Aec-5). Both together, in the sensitive genotype condition to AEC inhibition, showed only embryos sensitive to AEC (Fig. 1D) as observed for the Cat 100-1 inbred line (Fig. 1A). All other regenerated plants were sensitive to the inhibition caused by AEC.
Progenies derived from AEC-1 and AEC-5 resistant mutants and the AEC-10 and AEC-18 sensitive selected progenies, along with the Cat 100-1 inbred line (control), were used for soluble amino acid extraction and quantification by HPLC. Endosperms (30 per progeny) were individually analysed. It was observed that the AEC-10 sensitive progeny showed a 56% increase in the level of total soluble amino acids in comparison with the control, and also higher than the total soluble amino acid fraction of the resistant progenies analysed. Table 1 shows the results for soluble lysine levels. Although the absolute level of lysine was a little higher than in the control (0.24 and 0.19 µmol.g-1 endosperm in AEC-10 and Cat 100-1, respectively), its relative level was lower (3.78% of the total in the control and 3% of the total in AEC-10). For AEC-1 and AEC-5 the levels of soluble lysine were higher both in absolute (0.24 and 0.34 µmol.g-1 endosperm for AEC-1 and AEC-5, respectively) and relative (5.85% and 5.55% of the total for AEC-1 and AEC-5, respectively) levels.
Table 1. Soluble lysine of the endosperm of regenerated plans after
selection in medium containing AEC. Cat 100-1 represents the sensitive
control.
| Soluble lysine | ||
| Progeny | (µmolg-1 endosperm ± sd) | % of total |
| Cat 100-1 | 0.19±0.01 | 3.78 |
| AEC-1 | 0.24±0.07 | 5.85 |
| AEC-5 | 0.34±0.03 | 5.55 |
| AEC-10 | 0.24±0.04 | 3.00 |
| AEC-18 | 0.21±0.02 | 2.16 |
In maize, an accumulation of lysine in the endosperm has not been reported. This may be due to two main factors: first, for the lysine plus threonine resistant mutants, the DHDPS enzyme may still be sensitive to lysine blocking its synthesis at this regulation point of the aspartic acid pathway. Second, maize endosperms carry out a high rate of lysine degradation during seed development. This fact was first reported using [14C]-lysine (Sodek and Wilson, Arch. Biochem. Biophys. 140:25, 1970). Two other enzymes, lysine-a-ketoglutarate reductase (LKR) and saccharopine dehydrogenase (SDH), were detected and characterized in maize (Arruda et al., Plant Physiol. 69:988, 1982), and these enzymes are involved in the degradation of lysine. LKR converts lysine into saccharopine and is present at high activities in developing endosperms. This enzyme is also involved with the translocation of nitrogen to the seed (Arruda and Silva, Phytochemistry 22:2687, 1983). Two hypotheses should also be considered for the importance of lysine degradation as one of the main factors for the accumulation of lysine in seeds. Firstly, during seed development, translocation of lysine to the endosperm occurs in excess of 2-3 times the necessary level of lysine for protein synthesis (Arruda and Silva, Phytochemistry 18:409, 1979). The excess lysine translocated is degraded in normal endosperms, but this does not occur in mutants rich in lysine. Secondly, LKR shows very low activity in opaque2 endosperms, which are characterized by high levels of lysine (Arruda et al., MNL 58:50, 1984). This mutation does not affect SDH, therefore, LKR may be metabolising the excess lysine that could be accumulated in the resistant mutants. So, a further analysis for DHDPS in the AEC resistant mutants is being carried out to test these hypotheses.
Figure
1. Growth of excised embryos of regenerated plants in medium MO9-2
containing 0.25 mM AEC. (A) Cat 100-1 sensitive control, (B) regenerated
progeny AEC-1 showing dominant segregation for AEC resistance, (C) regenerated
progeny AEC-5 showing a recessive trait for AEC resistance and (D) hybrid
progeny (aec1 aec1 Aec5 Aec5) sensitive to AEC resistance. Bar =
5 mm.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}