QTL for drought responses in an F2 population
--Steve Quarrie, Claude Lebreton, Vesna Lazic-Jancic and Andrew Steed
We have been studying the physiological basis of differences in drought resistance amongst inbred maize lines for several years. Much of this work has been focused on the role of the stress hormone abscisic acid (ABA) in mediating stress responses. As ABA content appears to be inherited quantitatively we have started a program to map loci that regulate ABA production so that we can test for the presence of coincident QTL for a wide range of other traits associated with drought response.
We chose as our mapping population 81 F2 plants derived from a cross between two inbred lines that have been shown in previous work to differ in a wide range of responses to drought stress. In particular, they differ markedly in their leaf ABA contents, both under non-stressed and drought stressed conditions: F-2 (low-ABA, drought sensitive) and Polj17 (high-ABA, drought tolerant). Plants were grown in a soil glasshouse and sampled for leaf ABA content under non-stressed conditions at flowering time and then under mild drought stress after three weeks without water. Xylem sap was expressed from droughted leaves in a pressure chamber. At maturity plants were pulled out of the ground using a screw mechanism attached to a spring balance to measure the maximum force required to pull up the plant (root pulling force), and the number of roots in the root whorl at the base of the stem was counted. Leaf samples were also collected for anatomical measurements as the parents are known to differ significantly in xylem vessel dimensions and cuticular thickness.
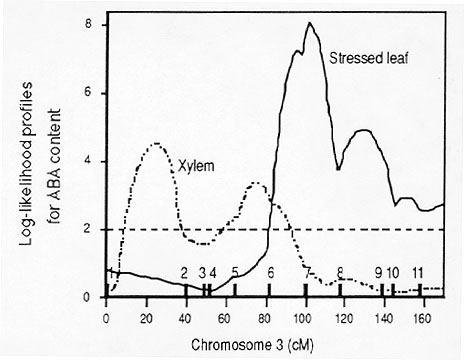
So far about 50 RFLP markers (mainly from the UMC collections) have been mapped, with an average of ten markers covering each of chromosomes 1, 3, 6 and 9. This allowed us to estimate the location on these chromosomes of QTL for the traits using MAPMAKER-QTL. Chromosome 1 showed evidence of QTL regulating ABA content in non-stressed and stressed leaves and in xylem sap. Chromosome 3 had major QTL for stressed leaf (maximum log-likelihood 8.0) and xylem sap (maximum log-likelihood 4.5) ABA contents, but on different regions of the chromosome (Fig. 1). No QTL for non-stressed leaf ABA content was found on this chromosome. Chromosome 6 had a significant QTL for xylem ABA content (maximum log-likelihood 2.6), but no evidence of any QTL for leaf ABA content.
QTL for stomatal conductance were widespread throughout the genome, and were consistently associated with QTL for leaf ABA content. The QTL for xylem ABA content on chromosome 6 was not associated with any effects on stomatal conductance, and it seems likely that in these plants the ABA content of the whole leaf had a greater role in the regulation of stomatal conductance than xylem ABA content. A major QTL determining root number was found on chromosome 8 and root pulling force was determined by loci on chromosomes 1, 3 and 8. In general, QTL for root pulling force coincided with QTL for stressed leaf ABA contents, high ABA content being associated with a high root pulling force. Thus, it is possible that ABA made in the leaves may have a role in stimulating root development in a drying soil.
We are currently adding further RFLP markers to the map and have collected seed from the self-pollinated F2 plants for further growth cabinet and field tests on the F3 generation to study the robustness of these drought-related QTL.
Figure
1. Log-likelihood profiles for stressed leaf and xylem sap ABA
contents for chromosome 3. Thick bars indicated the position of RFLP
markers on the chromosome. Markers: 1-umc121, 2-umc10,
3-umc102, 4-umc26, 5-bnl5.37, 6-umc60, 7-umc39,
8-umc16, 9-umc63, 10-umc96, 11-umc2a.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}