Mapping the centromere of chromosome 4 in maize using a telocentric
for 4S
--W. Lee, D. F. Weber, M. C. Schneerman, and G. Doyle
It is critical to correlate the maize genetic and cytological maps so that the rich array of maize cytogenetic variants can be used to manipulate its genome for use in molecular studies. One of the continuing goals of the Weber lab is to better correlate the genetic and cytological maps of maize. We are currently attempting to more accurately map the centromeres of each of the chromosomes on the maize RFLP genetic map. One approach we are using is with telocentric chromosomes.
Telocentrics have been recovered for several chromosome arms of the maize genome (Rhoades, Genetics 25:483-521, 1940; Doyle, MNL 62:49-50, 1988, Staub unpublished). Unfortunately, the telocentric for 5S that Rhoades (1940) recovered appears to have been lost (Dempsey, personal communication).
One of us (Doyle) has been analyzing progeny of trisomic maize plants that were heterozygous for a morphological marker locus on the trisomic chromosome. Genetic ratios were observed in certain of these crosses that indicated that a telocentric chromosome plus two normal homologs were present. A plant of this type is a telo-trisome. Here we report the use of a telocentric for the short arm of chromosome 4 (telo-4Sc) to more accurately map the position of the centromere of chromosome 4 on the maize RFLP map. One of us (Lee) analyzed telotrisome-4Sc plants cytologically and observed trivalents at diakinesis where one of the three members was smaller than the other two members. We are in the process of analyzing these at pachytene.
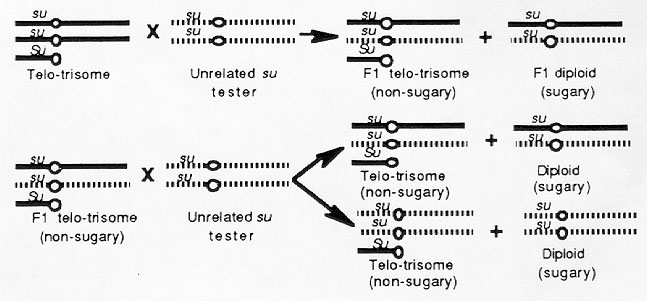
A telotrisomic stock for the short arm of chromosome 4 (telo-4Sc) had the recessive allele su on the normal chromosome 4's and Su on the telocentric chromosome. Part of the gametes produced by such a plant will contain a chromosome 4 (with su) and part will contain a chromosome 4 (with su) and a telo-4Sc (with Su). When a plant of this type is self-pollinated or testcrossed, two types of kernels are produced (not considering recombination between the Su locus on the telocentric and the centromere). Some are sugary, and these contain two normal 4s with su alleles. Others are non-sugary and contain two normal 4's plus the telo-4Sc. The progeny that contain the telocentric chromosome can be easily identified in this way.
Because this stock had been self-pollinated for several generations, the normal chromosome 4's and the telo-4Sc may be similar genetically. Therefore, the telotrisomic-4Sc stock was crossed as a female parent by an unrelated diploid tester stock that had the same recessive marker mutation (su), and the F1 non-sugary progeny were backcrossed as female parents to the same su tester stock. These crosses are diagrammed below. The chromosomes of the original stock are shown with solid lines and the chromosomes of the unrelated su tester are shown with broken lines.
When the F1 non-sugary progeny are backcrossed to the unrelated su tester, four types of progeny are produced. Half of the non-sugary kernels have both chromosome 4's from the tester and the other half have one from the original telotrisomic-4Sc stock and one from the su tester. Also, half of the sugary kernels have both chromosome 4's from the su tester and the other half have one from each parent.
For RFLP loci that have different alleles in the original telocentric-4Sc and su tester parents, the following relationships will exist. If the RFLP locus is located on the long arm of chromosome 4, half of the non-sugary backcross progeny will display both alleles and half will only display the allele that was present in the tester parent. However, if the locus is located on the short arm, both alleles will be displayed in all of the non-sugary progeny.
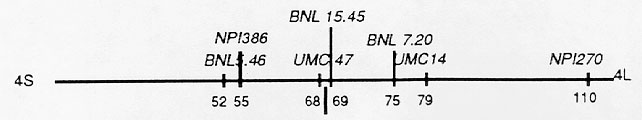
Leaf samples from the original telocentric-4Sc plant, the unrelated su tester, and non-sugary and sugary backcross progeny were harvested, freeze-dried, and DNAs were isolated from each plant type. We analyzed DNAs of the original telotrisomic-4Sc plant, the unrelated su tester, and ten telotrisomic (non-sugary) backcross progeny with a probe for RFLP locus BNL15.45. We found that the RFLP alleles for both the original telotrisome-4Sc and the tester parent were present in six of the F1 progeny and only the allele from the original telotrisome-4Sc was present in the other four F1s. A similar banding pattern was obtained for RFLP locus bnl7.20. These results indicate that these RFLP loci reside in the long arm of chromosome 4. When these same plants were explored with probes for RFLP loci bnl5.46, npi386, and umc47, both alleles were present in each of the F1s. Clearly, these RFLP loci are located in the short arm. Therefore, the centromere of chromosome 4 is located between RFLP loci bnl15.45 and umc47 as shown below:
Previously, we (Weber and Helentjaris, Genetics 121:583-590, 1989) explored B-A translocation hypoploids with RFLP probes. We were able to localize the centromere of chromosome 4 to a region between RFLP loci npi27, npi77, and npi95 (which mapped near each other in the short arm) and NPI250 in the long arm (a region spanning 26 map units). The current study localizes the position of the centromere to a region of 1 map unit. We will be exploring other telocentrics using the same experimental approach.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}