In view of the possible tetraploid nature of maize and consideration of its amphiploid derivative of n=5 of sorghum and n=5 of Coix by Anderson (Chronica Bot. 9:88-92, 1945) and Stonor and Anderson (Ann. Mo. Bot. Gard. 36:355-404), revisiting seems necessary with appropriate modification.
Tangible evidence in favour of x=5 as the basic chromosome number of the genus Zea has been provided from the secondary association in diploid maize (Chantekar, Cytologia 30:426-435, 1965), haploid maize (Majumdar and Sarkar, Cytologia 39:83-89, 1974; Ting, Maydica 30:161-169, 1985) and maize x teosinte hybrids by Ting (Bussey Inst. Harvard Univ. (monograph), p. 64, 1964), and in a series of papers by Molina and her associates (Cytologia 50:643-648, 1985; MNL 60:77-79, 1986, etc.). Electron micrographs of somatic metaphase chromosomes (Bennett, Kew Bot. Conf. II, pp. 71-79, 1983) also suggest the tetraploid nature of the maize genome. Buffering capacity and survival of monosomics (Weber, in W. F. Sheridan, ed., pp. 79-83, 1982) add further evidence for the amphiploid origin of maize.
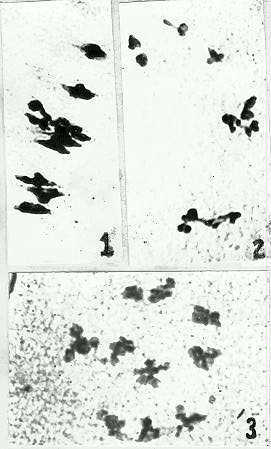
Our studies on secondary association in maize, teosinte and their hybrids have supported the amphiploid (2n=4x=20) nature of maize evolution. In maize (2n=20) and teosinte (2n=20) chromosome association has been observed in almost all the maize and teosinte hybrids. For example, maize x Zea diploperennis, maize x Z. luxurians, maize x Z. parviglumis and maize x Z. mexicana secondary association has been observed at different stages of meiosis in varying frequencies such as 5:5, 6:4, 3:3:2:2 and 2:2:2:2. The most frequent association at metaphase I was 3:3:2:2 (Fig. 2). It is interesting to note that the pattern of chromosome association is also retained in metaphase II (Figs. 1 and 3). Chromosome constellations observed in metaphase I are also faithfully reflected in metaphase II, which provides an opportunity to observe chromosome pairing and association etc. with more clarity than in metaphase I. Finally, it appears quite probable that the Zea species possess two homologous genomes, where pairing is restricted due to the presence of a Ph-like gene (Poggio et al., MNL 64:72, 1990).
Figure
1. Secondary association (3:3:2:2) at metaphase I in Nal-Tel x Zea
diploperennis.
Figure
2. Equatorial view of 3:3:2:2 constellation at metaphase Ii in Zea
diploperennis x CM105.
Figure
3. Polar view showing 3:3:2:2 constellation at metaphase II in CM105
x Zea diploperennis.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}