In the rhm tagging project, the mutation rates (susceptible Rhm to resistant rhm) for different elements usually ranged from 4 x 10-5 to 8 x 10-5 (see Chang and Peterson, accompanying report). The Cy population yielded a mutation rate of 4.17 x 10-5. No mutant was found for the T4-6(033-16)~c2-m1~Rhm population out of 47,391 seedlings screened. The two populations were combined into a heterozygote (crossed to increase vigor) and testcrossed by an rhm tester to develop screening seed. Screening of the testcross progeny from this heterozygote, however, yielded around 5% resistant mutants. The figure 5% was an estimate since no precise counting was made. This result certainly cannot be explained by random insertion of the element or natural mutation or both.
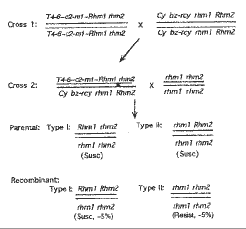
It is hypothesized that two closely linked recessive genes (rhm1 and rhm2) control resistance to Helminthosporium maydis (Bipolaris maydis), the causal fungus of southern leaf blight. The linkage distance is roughly 5% x 2 = 10%. Figure 1 is a diagram of the hypothesis proposed.
Figure 1. Model for resistance controlled by two linked recessive genes. T4-6, T4-6(033-16)
The two closely linked genes are temporarily designated rhm1 and rhm2, with no firm knowledge of the relationship of the two genes to the rhm gene proposed by Smith and Hooker (Crop Sci. 13:330-331, 1973). The dominant gene in T4-6(033-16)~c2-m1~Rhm is arbitrarily designated Rhm1 and that in the Cy line, Rhm2. According to this, the rhm tester will have two recessive genes. The two genes have coordinated effects so that only the genotype rhm1 rhm2//rhm1 rhm2 is resistant to the fungus. A dominant allele at either locus will abolish resistance. The cross described above between the two parents would have yielded a genotype with the two genes in repulsion phase, Rhm1 rhm2//rhm1 Rhm2. In the testcross progeny with the rhm tester, there are two types of parental genotypes which produce susceptible seedlings. There are two types of recombinants expected in the progeny, whose frequency depends on the linkage between the two genes. One of the two recombinants is rhm1 rhm2//rhm1 rhm2. It is this recombinant type that yielded the ~5% resistant "mutants". The other recombinant genotype Rhm1 Rhm2//rhm1 rhm2 is phenotypically indistinguishable from the majority (parental) seedlings. Therefore, the linkage distance between the two genes should be 5% x 2 = 10%.
A strategy has been developed to test this hypothesis. The rhm locus has been shown to be located at the end of 6S. Though an RFLP marker, umc85, was located close to the locus (Zaitlin et al., Genome 36:555-564, 1993), it was not determined whether this marker is proximal or distal to the locus. Thus a strategy based on RFLP analysis awaits a firm distal marker to the locus. The strategy presented here (Fig. 2) is a purely genetic one.
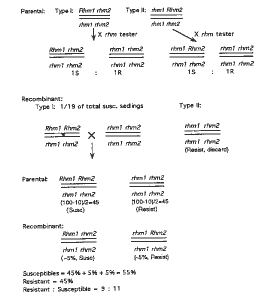
Figure 2. Test of hypothesis of resistance controlled by two linked recessive genes
1) The seed for screening (testcross progeny from the cross T4-6~c2-m1~Rhm1 rhm2//Cy bz-rcy rhm1 Rhm2 x rhm1 rhm2//rhm1 rhm2) is to be planted again. Among the progeny are the two types of nonrecombinant seedlings as well as the two types of recombinant seedlings. 2) Susceptible seedlings from lines that produce ~5% resistant individuals are to be transplanted and testcrossed by the same rhm tester. The two parental types will produce 1R : 1S seedlings since no crossover will yield a different phenotype. The recombinant type, however, will yield new recombinants. This will increase the proportion of susceptible seedlings by ~10%. Thus a new ratio 9R : 11S will result instead of 1R : 1S (10R : 10S).
The hypothesis of two linked rhm loci can be tested as outlined in Figure 2. The expected frequency of the susceptible recombinant class Rhm1 Rhm2//rhm1 rhm2 from cross 2 (Fig. 1) is 5% in the progeny. If the 5% resistant class is excluded, the frequency of susceptible recombinant class among all susceptible seedlings will be expected to be ~5% out of 95%, i.e., 1/19. This class can be recognized since it will produce a 9R : 11S distorted ratio. Using the formula given by Sedcole (Crop Sci. 17:667-668, 1977), the number of plants needed to be testcrossed is 67 in order to obtain at least one progeny yielding the distorted ratio at the 95% confidence level. Using the formula given by Clarke and Carbon (Cell 9:91-99, 1976), the number would be 55. Lastly, the testcross progeny will be tested for the ratio of resistant versus susceptible seedlings.
Chlorotic lesion resistance to southern leaf blight was first identified by Craig and Daniel-Kalio (Plant Dis. Rep. 53:134-136, 1968) in Nigeria. Craig and Fajemisin subsequently studied the inheritance of resistance. They reported (Craig and Fajemisin, Plant Dis. Rep. 53:742-743, 1969) that resistance was controlled by two closely linked recessive genes with a linkage of 16.83%. Using the material of these authors, Smith and Hooker (Crop Sci. 13:330-331, 1973) tested the inheritance of resistance on a large scale. From the data obtained, they concluded that resistance is controlled by a single recessive gene, designated rhm. However, Smith and Hooker used only one plant of the Nigerian material in the crosses. Utilization of a single plant could have excluded the second gene if the Nigerian material was not homogeneous. Later, Thompson and Bergquist (Crop Sci. 24:807-811, 1984) reported two independent recessive genes controlled seedling resistance, among other types. Holley and Goodman (Plant Dis. 73:562-564, 1989) reported new sources of resistance with more complicated inheritance patterns.
A few inferences can be drawn from this study. i) The data obtained in this experiment exclude the possibility of two independently segregating loci. The two genes involved in the two parental lines (cross 1, Fig. 1) were apparently linked. They behaved like the genes originally described by Craig and Fajemisin. ii) The two genes in this study interact in a coordinated manner similar to that proposed by Craig and Fajemisin and by Thompson and Bergquist. iii) One of the two genes could be the rhm gene proposed by Smith and Hooker, since the two parents, when separate, behaved as though only one gene were involved. iv) Although not possible to exclude, other epistatic effects, except the coordinated interaction described above, are not likely to have been a major force in determining resistance since the newly derived resistance type strongly resembled the rhm tester. This, at least, excludes the types of interaction observed by Holley and Goodman that resulted in intermediate levels of resistance.
How then to visualize the coordinated gene action that results in resistance with two genes? The most likely explanation is that one of the two genes is a copy of the other created by duplication. The dominant alleles are responsible for a gene product that permit the fungus to proliferate. The recessive alleles are null alleles. A dominant allele in either of the two copies of the gene will be able to make the gene product, and thus, will abolish resistance.
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}