The knotted1 (kn1) locus in maize is defined by a series of dominant alleles which cause sporadic localized outgrowths of tissue or knots on the lateral veins of the leaf blade. All of the existing alleles appear to be caused by insertion of transposable elements into introns of the gene except for Kn1-O, which is caused by a tandem duplication of the entire locus (reviewed by S. Hake, TIGS 8:109-114, 1992). The gene was cloned by transposon tagging (S. Hake, E. Vollbrecht and M. Freeling, EMBO J. 8:15-22, 1989) and shown to encode a homeodomain (E. Vollbrecht, B. Veit, N. Sinha and S. Hake, Nature 350:241-243, 1991). Analysis of the normal pattern of expression of kn1 indicates that the mRNA and the protein are abundant in apical meristems and immature, unexpanded axes of vegetative and floral shoots (L. Smith, B. Greene, B. Veit and S. Hake, Development 116:21-30, 1992; D. Jackson, B. Veit and S. Hake, in press). kn1 mRNA and KN1 protein are not detected in lateral organs of the plant and are apparently down-regulated even before the organ primordia are visible on the flanks of the meristem (Smith et al., 1992; Jackson et al., in press). Dominant Kn1 mutations cause the gene to be expressed ectopically in lateral veins of immature leaves (Smith et al., 1992). None of the dominant alleles appear to disrupt the normal pattern of expression; they merely add a new component.
The dominant mutations provide information about the role of the knotted1 gene outside of its normal context. In order to understand the role of kn1 in the meristem, loss of function alleles are needed. A small deletion of kn1, Def(Kn1)O, was isolated which removes the entire gene but not the closely linked loci alcohol dehydrogenase, adh1 (~1 cM), and lemon white (~1 cM) (J. Mathern and S. Hake, MNL 63:2, 1989). The deletion has no visible effects on the sporophyte as a heterozygote, but fails to pass through the male gametophyte. When Def(Kn1)O is uncovered in the progeny of crosses by the TB-1La translocation, the Def(Kn1)O-hypoploid embryo class displays an early embryonic lethal phenotype. We have recently found that at least one additional gene (knox3, a homeobox gene very similar in sequence and expression pattern to kn1) is missing in this deletion. Since it is unclear how many genes are missing in Def(Kn1)O, we cannot infer the phenotype of a loss of function allele of kn1 from the phenotype of the deletion.
We have used the chemical mutagen ethane methylsulfonic acid, EMS, to induce loss of function mutations, which can be selected by screening for the loss of the dominant Kn1-N2 phenotype. The Kn1-N2 allele is correlated with the presence of a receptor of Dotted element, rDt, in the fourth intron of the gene (N. Sinha, E. Vollbrecht and S. Hake, unpublished results) and is closely linked to the S allele of adh1. The phenotype is both highly expressive and fully penetrant in seedlings and it is characterized by wide, white veins and one to many knots on the first several leaves. Pollen was collected from plants heterozygous for Kn1-N2 and for Def(Kn1)O (which is never transmitted through the male gametophyte). The pollen was treated for 30 to 50 minutes in a 0.1% (v/v) emulsion of EMS in mineral oil according to the method of M. G. Neuffer (Mutagenesis in The Maize Handbook, eds. M. Freeling and V. Walbot, Springer-Verlag New York, Inc., pp. 212-219, 1993) and applied with a brush to silks of the inbred line B73 (Pioneer), which carries a normal allele of kn1 closely linked to the F allele of adh1. Seeds were collected and planted at high density and screened as seedlings for individuals without knots.
Of approximately 13,000 seedlings screened, 60 showed no sign of knots (this number included many self contaminants which might have been reduced had we detasseled the female parent). Various other mutations were observed in the population, including putative oil yellow seedlings, indicating the mutagenesis was effective. The EMS induced derivatives no longer expressing the knotted phenotype were selfed (where possible) and the progeny were examined for absence of knots, and tested for the closely linked adh1-S allele and for the presence of the rDt element associated with the Kn1-N2 allele in order to weed out normal individuals resulting from pollen contaminants. Ten families carried the proper markers and continued to show no sign of knots.
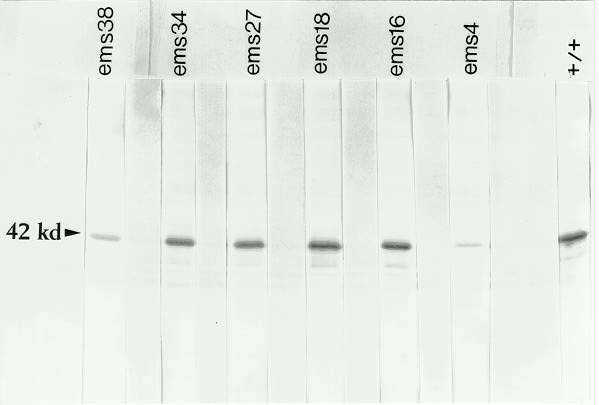
As a preliminary step toward characterizing the changes at the kn1 locus resulting from EMS mutagenesis, we looked for changes in the KN1 protein produced by these Kn1-N2 derivatives on Western blots. Homozygous adh1-S seeds from each were planted, and seedlings were harvested at 3 to 5 weeks after planting. Leaves longer than 5mm were removed and the meristem and a few millimeters of ground tissue were harvested and proteins extracted. Western blots were made and the KN1 protein was visualized with affinity purified polyclonal antibody specific to KN1 (as in Smith et al., 1992). Equal quantities of protein were loaded in each lane and protein from B73 meristems was run for comparison. The results for 6 out of 10 of the derivatives are shown in Figure 1. Each derivative family produces at least some KN1 protein, and no novel proteins reacting with the anti-KN1 antibody were observed. Four derivative families produce normal levels of KN1 protein (shown in Figure 1: ems34, ems27, ems18 and ems16), while six show significant reductions in the levels of KN1 protein, possibly due to a point mutation rendering the protein unstable (two of these shown in Figure 1: ems38 and ems4). Several of the derivatives still show some level of ectopic expression in young leaf tissue (data not shown). In each case, however, the levels appear lower than in leaf tissue from a homozygous Kn1-N2 mutant. All of the homozygous adh1-S progeny of the derivatives appear normal as seedlings, including the two with the greatest reductions in KN1 protein levels, ems38 and ems4. A thorough examination of adult phenotypes is in progress. So far, homozygotes from one derivative (R848-6) appear to have markedly reduced seed set when compared to heterozygous sibs.
Our efforts to generate loss of function alleles at the knotted1 locus by EMS mutagenesis of a mutant allele have yielded several new alleles that no longer condition the Knotted phenotype. Since all of them make at least some KN1 protein, we may not yet have isolated a complete null allele. The isolation of derivatives that have significantly reduced levels of the KN1 protein suggests we have isolated events that reduce expression below a threshold necessary to induce knot formation or that alter the stability of KN1 protein. Derivatives with levels of protein comparable to normal suggest qualitative alterations of the KN1 protein that may impair its function or cause loss of its expression in the cells necessary to cause knot formation. Further characterization of the specific defects may point toward significant elements of the protein or promoter of the maize homeobox gene, kn1.
Figure
1. Western blot with anti-KN1 antibody on proteins extracted from individuals
homozygous for six different EMS-induced derivatives of Kn1-N2 that
no longer condition the Kn1 mutant phenotype, and a wild-type control
(right end). Methods and materials as described in Smith et al. (Development
116:21-30, 1992).
Return to the MNL 68 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}