Early in development maize embryos acquire the capacity to germinate or to mature. During maturation phase embryos accumulate nutritional reserves, gain tolerance to desiccation and become metabolically quiescent. One approach we have taken to increase our understanding of maturation has been to analyze embryo mutants that are blocked from normal passage through this phase. We have been studying a group of defective kernel (dek) mutants which were reported by Neuffer et al. (1986) to undergo morphogenetic blocks after forming only two or three leaf primordia. This stage of shoot development corresponds to the time when wildtype embryos enter maturation phase.
We have been investigating the developmental potential of this class of dek mutant by characterizing their growth, morphogenesis, accumulation of maturation products, and ability to germinate over the course of ontogeny. We report here the results from our descriptive and experimental studies of two dek mutants, cp*-1311C and cp*-1399A, kindly given to us by Sheridan, WF and Neuffer, MG.
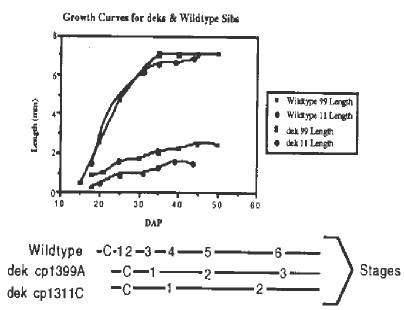
The first question we asked of these mutants was whether or not the mutations represented blocks to continued development. We compared growth (size, fresh and dry weights) and morphological stages of mutant embryos to that of their wildtype counterparts at various time points during embryogenesis. Cumulative growth profiles of deks cp*-1311C and cp*-1399A and their wildtype sibs (see figure) show that these embryos do not become developmentally arrested. Instead, dek embryo growth and morphogenesis is slow and steady from early in development until metabolic quiescence near full term, rather than arrested. We suspect that these mutants are altered in some component of nutrient uptake or in a metabolic pathway rather than in the maturation process.
These mutants reach morphological stages associated with early maturation events in wildtype embryos, but their timetables of development are not synchronized with the changing maternal environment. We suspected that this situation would allow maternal cues for development to affect the mutant embryos at age- rather than stage-appropriate times, thereby allowing us to distinguish between environmental and seed-autonomous cues. To determine if and when dek embryos matured and whether biochemical differentiation remained coupled to growth and morphogenesis, we looked for the accumulation of pigments, storage macromolecules and LEA proteins (proteins associated with dehydration). We compared the results from mutant and wildtype embryos, matched by either morphological stage or age (days after pollination). In both dek cp*-1311C and cp*-1399A embryos, carotenoids, anthocyanins and the major embryo storage globulin protein, GLB1, accumulated properly according to the size and morphological stage of the mutant embryos. Linkage to morphological stage suggests that cues for the accumulation of pigments and storage proteins are seed-autonomous. In contrast, both dek mutants accumulated the LEA group 3 protein, MLG3, at the same time and in the same amount as their wildtype siblings. The uncoupling of MLG3 accumulation from dek embryo morphogenesis suggests that the accumulation of MLG3 is due to a maternal cue.
The acquisition of desiccation tolerance and ability to germinate are additional features of the maturation process. dek cp*-1311C and cp*-1399A embryos were able to germinate precociously, from 18 days after pollination until full ear maturity (albeit less vigorously than wildtypes) when embryos were cultured in a hormone-free germination media (GM). However, after 60 DAP they were unable to germinate, either as whole kernels or dissected embryos. To test whether deks lose their ability to germinate because of an intolerance to desiccation and subsequent imbibition, we used fluoroscene diacetate (0.1%) as a vital stain on ear mature dek, wildtype (positive control) and heat-killed wildtype embryos (negative control). The embryos were soaked in water overnight at room temperature, sectioned longitudinally and then viewed under ultraviolet light. dek cp*-1399A scutella fluoresced as brightly as the wildtype, while dek cp*-1311C scutella fluoresced very little. Desiccation tolerance is gained by wildtype embryos by stage 4 (ca. 30 DAP). Since the deks (especially cp*-1399A) can withstand maturation drying, they must be desiccation tolerant prematurely with respect to their terminal morphological stages.
To further assess the physiological age of mutant embryos, we studied their behavior in embryo-culture experiments. Immature and mature mutant and wildtype embryos were cultured in GM, or GM plus abscisic acid (ABA) or osmoticum. We have previously shown that wildtype embryos have an age-dependent sensitivity to these cues for maturation. Wildtype embryos at stages 2 and 3 (18-22 DAP) germinate readily in GM alone but are inhibited from germination by both 10µM ABA and 20% sucrose (osmoticum). By 50 DAP the germination of wildtype embryos is unaffected by ABA and only slightly slowed by osmoticum. In contrast, dek embryos at 50 DAP (stage 2 or 3) respond to the culture treatments in a stage- rather than age-appropriate fashion -- i.e., both ABA and osmoticum inhibit germination. Therefore, before the dek embryos completely lose their ability to germinate following desiccation, their response to culture treatments is stage-appropriate.
Figure.
Growth Curves for deks & Wildtype Sibs
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}