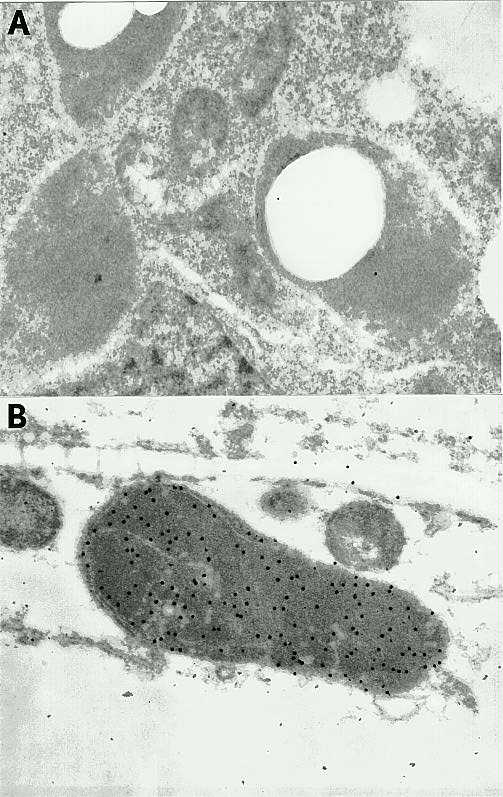
The seedling parts (primary and secondary roots and coleoptile node, which contains the shoot apex and primordial leaves) that are known to be rich in enzyme activity were fixed, sectioned, and incubated with anti-b-glucosidase sera and protein A-conjugated to 20nm gold particles, respectively. Preimmune and nonimmune sera served as controls. The examination of the distribution and intensity of gold labelling revealed that the label was found exclusively in plastids in root cells and proplastids and immature chloroplasts in shoot cells (Figure 1, B). There was no gold labelling in sections incubated with pre-immune serum (Figure 1, A) while there was fairly intense labelling in all sections incubated with immune serum indicating that labeling was specific for the enzyme. At this point it appears that the enzyme is localized to the stroma portion of the plastid; but this conclusion needs to be confirmed by further studies. The cell wall, which is the site of b-glucosidase localization in legumes and other dicots, does not seem to have any b-glucosidase in maize. Sorghum (Thayer and Conn, 1981), oats (Nisius, 1988) and rice (Muslim and Esen, unpublished) b-glucosidases have been also localized to the plastid, suggesting that this organelle is the site of the enzyme in all grasses, if not in monocots.
Figure 1. Immunocytochemical localizations of maize b-glucosidase to proplastids in shoots. A. A cell with several plastid sections showing no labelling after immunostaining with the preimmune serum. B. Section of proplastids in cells from the shoot apex region showing intense gold labelling (20nm) after incubation with anti-b-glucosidase antiserum and gold conjugation protein A.
The localization of maize b-glucosidase to the plastid by immunocytochemistry was also confirmed by two related approaches. First, etioplasts and chloroplast were isolated from young seedling parts (shoots including primordial leaves, and primary roots) and stained histochemically by infusing the substrate 6-bromo-2-naphthyl-b-D-glucoside plus a coupling dye or X-glu. Only the plastids showed staining (b-glucosidase activity). The same results were obtained when isolated plastids were incubated with substrates. Second, young seedling parts were free-hand sectioned and incubated with the same substrate and coupling dye combination as that used for plastids. Microscopic examinations showed clearly that the enzyme activity (i.e., staining) was associated with discrete organelles, identified as plastids. Histochemical staining was judged to be specific for b-glucosidase because it was not observed in the presence of such known inhibitors of b-glucosidase activity as b-gluconolactone, silver or mercury ions. Moreover, in the case of primary roots, histochemical staining was highest in the root cap and in the zone of elongation including root hairs. In fact, root hairs had discrete packages of b-glucosidase-positive organelles (plastids) occurring at intervals along their entire length as well as their tips. The implications of these findings are profound because they suggest that (1) the precursor enzyme monomer will include a transit sequence to target it to the plastid, and (2) the plastid is the organelle in which the enzyme performs its catalytic functions and also a likely site for the enzyme's physiological substrates. Alternatively, the plastid might be the site for sequestering and storing the enzyme.
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}