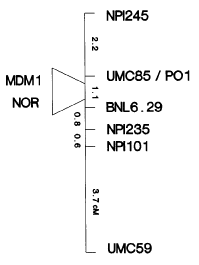
The satellite region distal to the NOR on the short arm of chromosome six contains resistance genes to two potyviruses, Maize Dwarf Mosaic Virus (Mdm1) and Wheat Streak Mosaic Virus (Wsm1), and a recessive source of resistance to the fungal pathogen Cochliobolus heterostrophus race O (rhm1).
Maize Dwarf Mosaic Virus. Several unlinked loci are involved in resistance in maize to MDMV. Presently, all characterized sources of resistance to MDMV have involved the Mdm1 allele on the short arm of chromosome 6. The short arm of chromosome 6 is also responsible for resistance to all known strains of MDMV. McMullen and Louie (MPMI 2:309, 1989) initially placed mdm1 in the interval between the distal marker, umc85, and the proximal marker, bnl6.29. A F2 population was generated using the MDMV resistant inbred Pa405 and a susceptible B73 inbred homozygous recessive for polymitotic1 (po1 mdm1) to further characterize the map position of mdm1. F2 progenies were planted in the summer of 1991 disease nursery and inoculated with MDMV strain A (MDMV-Å). DNA was prepared according to McMullen and Louie (1989) from whorl tissue of each F2 individual and genotype using RFLP markers on the short arm of chromosome 6. Plants were rated for resistance to MDMV-A three times throughout the growing season and scored for fertility. During the winter of 1991, field scores for resistance of some individuals were also confirmed by screening F3 progeny in the greenhouse.
The F2 mapping data cleared up several ambiguities in the reported map order of RFLP markers on chromosome 6S. The position of the NOR was found to map within the umc85-bnl6.29 interval (Figure 1). The NOR probe, pZMS1, containing the 3.0kb spacer region, detects an EcoRI polymorphism in about 10% of the repeat units in Pa405. Because of the difficulty in scoring the heterozygotes (5% versus 10%) the NOR was scored as a dominant marker. Data from a Pa405/y M14 x y M14 backcross and analysis of F2 recombinants placed the NOR proximal to umc85 and distal to bnl6.29a (Table 1). The po1 locus was found to be tightly linked to umc85 and we have yet to identify a recombinant between these two loci. Mapping of mdm1 in our F2 population agreed with the initial placement of mdm1 within the umc85-bnl6.29 interval distal to the NOR (Figure 1). Our best estimate would place mdm1 0.5cM proximal to umc85. One difference between other 6S RFLP maps and our mapping data is that we have less recombination across 6S. Our F2 recombination data agree with the map distances produced from a Pa405/y M14 x y M14 backcross.
Table 1. Number of recombinant chromosomes between po1, mdm1,
NOR, and RFLP markers on the short arm of chromosome 6.
| Interval | #Recomb. | Interval | #Recomb. |
|
|
|||
| npi245--umc85/po1 |
|
bnl6.29--npi235 |
|
| umc85 /po1--mdm1 | 2 | npi235--npi101 | 3 |
| mdm1--NOR | 1 | npi101--umc59 | 13 |
|
|
|||
| umc85--mdm 1 | 2 | bnl6.29-npi101 | 3 |
| mdm1--NOR | 1 | npi101--y1 | 16 |
| NOR--bnl6.29 | 3 | ||
Wheat Streak Mosaic Virus. Although WSMV is an economically important pathogen of wheat, maize acts as an oversummering host. Only a few maize genotypes are susceptible to WSMV, and these lines are also susceptible to MDMV. These two viruses share approximately 38% protein coat similarity, but WSMV and MDMV differ in both symptoms produced in susceptible host and vector relationships. Resistance in maize to WSMV was first genetically defined by M. D. McMullen and R. Louie (Phytopath. 81:624, 1991) and appears to be conferred by at least two dominant genes, Wsm1 and Wsm2. In crosses between Pa405 and the WSMV susceptible inbred Oh28, either gene is sufficient to confer resistance to WSMV. wsm1 has been mapped to the short arm of chromosome 6 and is tightly linked to po1/umc85 (data not shown). The mapping of wsm1 near mdm1 shows that these genes are either allelic or are tightly linked.
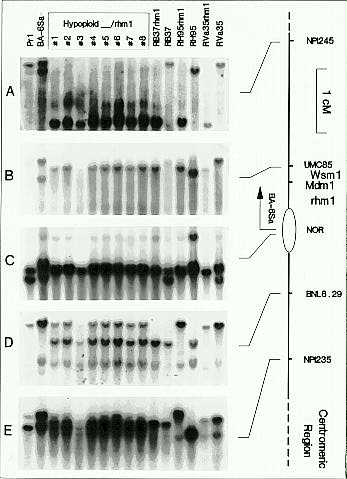
Cochliobolus heterostrophus. A recessive chlorotic-lesion type resistance to C. heterostrophus, causal agent of the southern corn leaf blight, was first identified in a Nigerian inbred in 1968 (J. Craig and J. M. Fajemisin, Plant Dis. Reptr. 53:742, 1969). D. Smith and A. Hooker proposed the symbol rhm1 for the recessive chlorotic-lesion resistance and placed this locus on chromosome 6 using a series of y1-marked reciprocal translocations (Crop Sci. 13:330, 1973). Zaitlin and co-workers (Genome, in press, 1993) have mapped rhm1 to the short arm of chromosome 6, near umc85. We are interested in determining the location of rhm1 in relation to mdm1 and the NOR. Two approaches were used to localize rhm1, B-A translocations which are used to identify loci distal to the A-chromosome breakpoint and near-isogenic line analysis (NIL's), which defines the chromosome segment retained during introgression of rhm1. The B-A translocation TB-6Sa was crossed as a male onto the inbred RB37 rhm1 and the F1 progeny were screened for resistance to C. heterostrophus. If rhm1 is distal to the NOR, resistant F1 progenies could result from fertilization of the egg (6, rhm1) by a sperm nucleus hypoploid (6B) for the region of chromosome six distal to the reported breakpoint of TB-6Sa in the NOR. Resistant progenies were recovered in the F1 and subjected to RFLP analysis. The markers proximal to the TB-6Sa breakpoint (bnl6.29a and npi235) detected both the RB37 rhm1 and the TB-6Sa alleles in the resistant hypoploid progeny, verifying that the TB-6Sa breakpoint and the rhm1 locus were distal to bnl6.29a (Figure 2, panel D and E, respectively). RFLP markers distal to the TB-6Sa breakpoint (npi245, umc85, and the EcoRI polymorphism detected with the rDNA spacer probe) were hemizygous for the RB37 rhm1 allele in all of the hypoploid progeny (Figure 2, panels A, B, and C, respectively). The B-A translocation data indicate that the rhm1 locus maps distal to the breakpoint in the NOR heterochromatin.
Near-isogenic line analysis of three rhm1 conversions (RB37 rhm1,
RH95 rhm1, and RVa35 rhm1) suggests that the chromosome segment
retained from the Nigerian inbred 024-2-4, through at least 6 backcross
generations, extends from the EcoRI polymorphism in the rDNA spacer
region through npi245. The same RFLP allele is present in all rhm1
converted lines when probed with npi245, umc85, and the rDNA
spacer probe (Figure 2, panels A, B, and C, respectively). All recurrent
parents were polymorphic with their respective converted rhm1 line
for these RFLP markers. The RB37 rhm1 and RH95 rhm1 conversions
matched their respective recurrent parents at the bnl6.29 and npi235
loci, indicating that the introgressed chromosome segment from the Nigerian
inbred 024-2-4 was entirely distal to bnl6.29 (Figure 2, panels
D and E, respectively). However, the introgressed segment in RH95 rhm1
may extend through bnl6.29 and npi235. Inoculation of the
three rhm1 conversion lines with MDMV-Å, -B, and WSMV indicates
that the introgressed segment from the original Nigerian source of rhm1
carried both mdm1 and wsm1 as well as rhm1. We are
currently crossing Pa405 and RH95 rhm1 to map mdm1 and wsm1
in relation to rhm1.
Figure 1. RFLP linkage map of the short arm of chromosome 6.
Figure
2. B-A translocation and near isogenic line analysis of rhm1
conversions. DNA's represented in lanes 1 through 16 are: Pr1 and TB-6Sa/Pr1
(lanes 1 and 2), hypoploid F1 progenies of RB37 rhm1/TB-6Sa) lanes
3 through 10), and the rhm1 conversion lines with the corresponding
recurrent parents (lanes 11 through 16). Panels A through E represent the
same hybridization membrane probed with npi245 (panel A), umc85
(panel B), pZMS1 (panel C), bnl6.29a (panel D), and npi235
(panel E).
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}