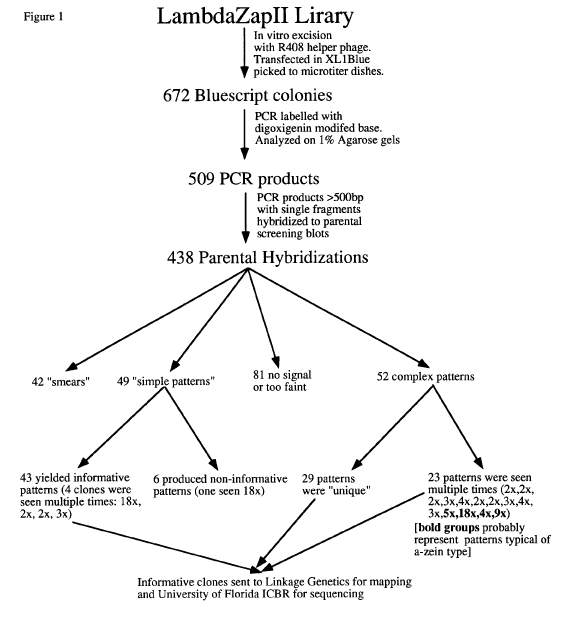
In our undertaking to enlarge the maize molecular marker map, we have begun initial efforts to both map and sequence randomly chosen cDNA clones. Two different libraries have been investigated. The first was constructed in LambdaZapII with NotI-EcoRI linkers ligated to ds cDNAs prepared from 14 DAP sweet corn endosperms. No size selection was incorporated prior to ligation to the EcoRI site of the phage arms. Bluescript phagemids were excised from the library using R408 helper phage, transfected into E. coli XL1Blue cells, and plated out on selective media. 672 colonies were picked into microtiter dishes and screened. Cultures of each colony were used as template in PCR labelling reactions containing a digoxigenin-modified nucleotide. The probes were hybridized to Southern blots and detected using AMPPD chemiluminesence activated by an Ab-conjugate of alkaline phosphatase. The Brookhaven recombinant inbred collection was used for assigning map positions for all polymorphic loci detected by these clones.
The results of this initial screen are summarized in Figure
1. For a number of reasons, many clones from this library proved unsuitable
at different steps in the process and had to be discarded. The resulting
informative clones (both complex and simple) were sent to Linkage Genetics
(Salt Lake City, Utah) for
mapping with the RI progeny. The mapping data yielded 99 probes that
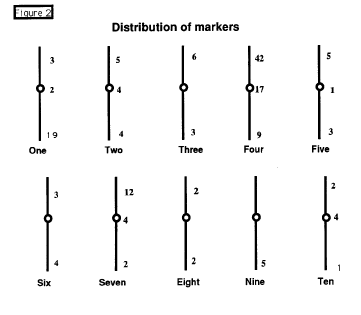
mapped to 161 loci. These probes are found throughout the genome (see Figure
2) with an asymmetric distribution. Particularly noteworthy is the
large number of markers mapped to the short arm of chromosome 4, many of
which we have determined to represent various zein genes. This should not
be surprising as zeins comprise a large percentage of expressed genes in
developing seeds. Interestingly most of these zein clones are found in
a large "inverted" array comprising most of 4S. Prior to this study there
were very few RFLP markers on this chromosome arm, which suggests several
interesting possibilities for consideration. Most RFLP markers in the past
have utilized genomic clones from PstI libraries as a selection
for low-copy-number sequences. That some areas of the genome are underrepresented
by this general pool could suggest that methylation occurs non-randomly
in maize. Alternatively different areas of the genome may contain different
levels of unique sequences. In any case, the use of randomly chosen cDNAs
prepared from different tissues may facilitate closing some of the "gaps"
in the current maize molecular marker map.
Many of these clones were also sent to the University of Florida (R. Ferl, R) for sequencing. Very few have been sequenced to date but it is our intention to both map and sequence all of these clones in order to attempt to assign both functions and genome locations to these cDNAs. One clone was found that possessed a short but significant homology to bt1 and mapped to at least five different chromosomal locations, suggesting that this strategy should produce some interesting insights at the same time we are enlarging the maize marker map. While our success rate from start to finish with this library has been lower than we would like, we have been much more successful with a library prepared by Stratagene of maize leaves, and many of these clones previously have been sequenced by C. Baysdorfer, C at California State University. >90% of the clones picked from this library amplify and we have been able to detect genomic polymorphisms for almost all of these. The difference we believe lies in the construction of the libraries. The first library was constructed without imposing any size selection upon the inserts and many have been in the range of only 150-200bp, which is too small to produce an effective hybridization signal or meaningful sequence comparison to existing databases. The latter utilized an oligo-dT-primer-linker and a size selection which resulted in most of the randomly-chosen clones being in the +500bp range. We believe this fact explains most of the differences we have observed in our success and would urge others to utilize both steps in constructing their own cDNA libraries, no matter what their desired purpose.
For future use we are constructing cDNA libraries using RNA isolated from both etiolated seedlings and developing seeds. However three principal differences in the construction will be used: 1) LambdaZipLox (Gibco BRL) will be used as a cloning vector which possesses the site-specific Cre-lox recombinational machinery to provide automatic in vivo subcloning, 2) directional libraries will be constructed utilizing NotI-oligo-dT primers, SalI adapters and SalI-NotI digested arms, and 3) the cDNA will be size-selected using a sephadex column before ligating it to the vector arms. Once this library is prepared clones will be isolated from it in three classes as either "abundant and common between tissues", "abundant and specific to the tissue of origin", and as "rare" using subtraction and screening strategies. We will then proceed with sequencing and mapping of this set of clones to determine how much more efficient this process is with this strategy.
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}