Maize is a chilling-sensitive plant: temperatures below 15 C suppress growth and prolonged exposure to temperatures of 10 C or less will kill seedlings. We previously reported that maize seedlings subjected to a chilling stress at 15 C for up to a few days accumulate Adh1 mRNA and ADH1 protein and enzymatic activity (Christie et al., Plant Physiol. 95:699-705, 1991); within 2 days of return to normal growing conditions, Adh1 mRNA and protein levels return to pre-stress levels. Because ADH1 can partially compensate the cell for a loss of respiration during hypoxia, we hypothesized that the rapid appearance and strong induction of ADH1 activity indicated suppression of mitochondrial respiratory activity during chilling treatment. We also reasoned that plants lacking ADH1 or doubly null (ADH1- ADH2-) would be more severely affected by cold treatment than normal seedlings.
To compare the chilling sensitivity of B37N and ADH1- or ADH1- ADH2- plants, five seedling growth experiments were conducted under slightly different conditions. Plants were germinated for 7 days in growth chambers at 25 C-30 C with 12-16 hours of light/24 hours. For chilling treatment, trays of seedlings were switched to a growth chamber kept at 10-15 C with the same light regime as the control seedlings remaining at 25-27 C. Visual inspection of B37N seedlings after 1-7 days at 15 C indicated that anthocyanin pigmentation was strongly induced in the leaf sheaths compared to the control seedlings, growth was almost completely suppressed, and the seedling leaf tips were desiccated. The seedlings survived upon return to control conditions. Up to 50% mortality of B37N seedlings, measured as a failure to grow when returned to control conditions, was achieved after 14-18 days of chilling stress. In contrast, the ADH1- and ADH1- ADH2- seedlings were more sensitive to chilling stress, with up to 50% mortality after 4 days of cool temperature treatment. The seedlings exhibited water-logging followed by desiccation of the leaf tips to a greater extent than did chilled B37N seedlings. The control ADH-deficient plants grew more slowly than did control B37N seedlings, so direct comparisons of these genotypes is not possible, however, wet and dry weight accumulation was decreased more in the chilled ADH-deficient lines compared to their control than was the case for B37N. These growth experiments substantiate the hypothesis that ADH expression is likely to be important for long-term survival during chilling stress.
The hypothesis that mitochondrial respiration would be affected by chilling stress was tested by measuring oxygen consumption. The measurements were made in 0.1M phosphate buffer, pH 6.3, or in this buffer with addition of either 10mM SHAM or 0.5mM KCN to selectively inhibit alternative oxidase or cytochrome oxidase activity, respectively. Seedlings were grown for 10 days at 31-32 C with 12 hr 85uE m-2/12 hr dark; control seedlings remained in these conditions. For chilling treatment, trays of seedlings were transferred to 15 C day/10 C night with the same light/dark regime. In each experiment, four replicate samples were examined for each condition. Each experiment was repeated two or three times; the results from one experiment are reported here.
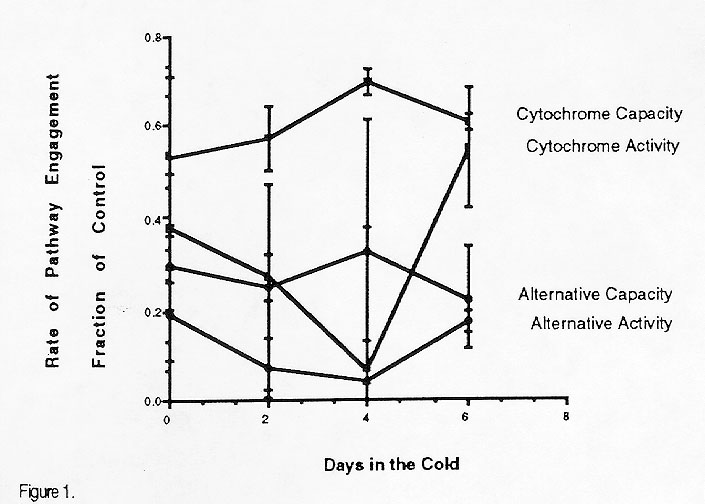
Each sample consisted of root tissue from one seedling; 0.1g of root tissue was sliced into 0.5cm sections in buffer just prior to each measurement. Oxidase activity and oxidase capacity were determined using standard methods (Moeller et al., Physiol. Plant. 72:642-649, 1988) in a Hansatech oxygen electrode at 35 C; an A/D board in an IBM-PC transformed the electrode output into a data file. Oxidase capacity is defined as the maximum level of pathway engagement measured when a competing pathway that uses substrate for the same function is fully inhibited. Oxidase activity is a measure of the degree of pathway engagement when competing pathways are both active. With B37N (see Figure 1), about twice as much oxygen consumption occurs through the cytochrome oxidase pathway as in the alternative oxidase pathway in roots of 10 day-old seedlings. Note that the 4 control samples exhibited virtually identical respiratory properties in this (and other) experiments. During cold treatment, however, root-to-root variation in respiration can be very large. This is particularly striking for cytochrome oxidase activity after 4 days of cold treatment. We interpret this variation as reflecting true physiological differences among the seedlings; that is, in some seedlings cytochrome oxidase activity is greatly depressed by cold treatment. We do not understand the origin of these differences between seedlings; only seedlings of similar size (developmental age) were selected for measurement.
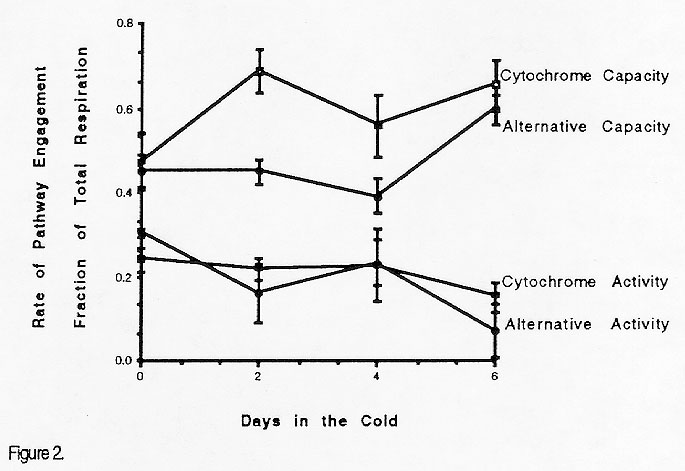
Compared to B37N seedling roots, the 10 day-old ADH1- ADH2- double null roots had much lower cytochrome oxidase activity (Figure 2). Both the activities and capacities of the two oxidases were similar in the double nulls. The ADH- material is in a different nuclear background than B37N, consequently either this difference or the absence of ADH could impact oxidase content. Similar to the B37N seedlings, the ADH double null seedlings also showed greater root-to-root variation in cold-stressed materials than in the ambient control populations (Figure 2).
Contradictory to our initial hypothesis neither oxidase capacity nor activity was significantly affected by chilling stress. In the B37N seedlings with normal levels of ADH, cytochrome oxidase and alternative oxidase capacities and activities remained relatively constant through 6 days of cold treatment. Increases in alternative oxidase have been measured after chilling treatment of other tissues (such as sliced potato tubers), however, we find no evidence for preferential use of alternative oxidase during chilling treatment of B37N seedlings. In the ADH double nulls, the two oxidases also remain relatively constant in both activity and capacity. There is, however, a significant increase in alternative oxidase capacity from after 6 days in the cold. A substantial change in capacity probably reflects increased accumulation of alternative oxidase protein after prolonged chilling treatment.
In summary we have found that respiratory activity and capacity are relatively insensitive to chilling stress in two genotypes of maize and are certainly not rapidly modulated by chilling. Both alternative oxidase and cytochrome oxidase activities and capacities remain relatively constant through 6 days of chilling treatment. Thus, it is unlikely that the rapid induction of ADH activity in normal seedlings is in direct response to a lack of mitochondrial respiration. ADH activity may be required to modulate cytoplasmic pools of key metabolites during chilling stress. This is a different role than observed during oxygen deprivation where maintenance of glycolysis and ATP production by production of catalytic amounts of NAD+ is the key contribution of ADH.
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}