Earlier studies had identified isolates of the inbred line A188 carrying a heritable activity that increased the frequency of somatic mutations of several maize genes (Das et al., PNAS 87:7809, 1990, Das and Messing, Methods in Molecular Genetics, K. W. Adolph, ed., Academic Press, Orlando, In Press). Somatic mutations of the P gene in these A188 isolates and their derivatives were recovered as colorless pericarp sectors on the uniform red background in P-rr/P-ww plants. Mutations causing a pericarp sector can be transmitted to the underlying kernel at a maximal frequency of 50% if the sector covers the germinal half or more of a kernel (Anderson, EG and Brink, RA, Am. J. Bot. 39:637, 1952). Among progeny ears from eight kernels bearing such sectors, two showed a patterned or variegated phenotype in pericarp. This phenotype was not seen among >6,000 ears screened over the course of these experiments, indicating that transmission of the mutation causing the sector was responsible for the new phenotype in the two cases.
Inheritance of the patterned pericarp phenotype has been studied over five additional generations of one isolate, and two of the other. Genetic analysis of three generations of progeny from one sectored kernel and two generations from the other is presented in Fig. 1. The sectored kernels were from ears obtained by growing P-rr/P-ww plants and their F2 progeny in an isolation plot. Both kernels evidently received P-ww from the male parent, as judged by segregation in the succeeding generation (Fig. 1). Segregation of the patterned phenotype was consistent with the phenotype being conditioned by an allele of P, designated P-pr for patterned pericarp and red cob. Pigmentation conditioned by this allele in pericarp, cob and tassel glume edges was highly variable, but variegation could be detected only in pericarp. Pigmentation in pericarp varied from color only at the silk attachment point to almost full color, and cob color varied from almost colorless to red. Homozygotes among sibling progeny displayed less pigmentation in pericarp than heterozygotes. But this was insufficient to account for the extent of variation in pericarp color, and it is not yet clear if the additional variation has a genetic basis.
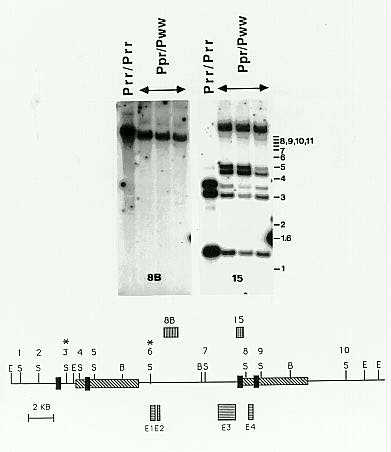
Molecular analysis using two probes derived from the P gene (kindly provided by Dr. Thomas Peterson, T; Lechelt et al., MGG 219:225, 1989; Grotewold et al., PNAS 88:4587, 1991), and 27 methylation-insensitive restriction enzymes showed no differences between P-pr and P-rr. This indicated that no major insertion, deletion or rearrangement of the P gene was associated with the mutation to P-pr. However, eight restriction enzymes sensitive to cytosine methylation showed that the P gene in leaf DNA from plants carrying P-pr was hypermethylated relative to P-rr. Representative data with SalI are shown in Fig. 2. Increased methylation was detected with probe 15, but not with probe 8B (Fig. 2). The location of the two probes showed that 5' and 3' flanking sequences of the gene were hypermethylated, but not the intervening region covering most of the transcribed region. In particular, the 5' region is located >5kb away from the 5' end of the functional transcript; Ac insertions in this region affect phenotype (Moreno et al., Genetics 131:939, 1991), indicating that it carries determinants of gene function.
To test if methylation correlated with phenotype, leaf DNA from 38 plants of #6601, 40 of #6671, 24 of #6672 and #6673, 14 of #6701 and 10 of #6702 was analyzed on Southern blots probed with probe 15 (family numbers are defined in Fig. 1). A methylation-insensitive restriction enzyme was used to identify P alleles, and SalI was used to determine their methylation status. Among these 126 samples, 90 carried a P-rr-like gene as identified by the first enzyme. All 90 showed at least one of the higher MW SalI bands corresponding to hypermethylation. No methylation changes were detected for P-ww. All but one of the 90 had patterned pericarp. The ear from the exceptional plant had fully red pericarp. This was the least hypermethylated sample, with only one of the two higher MW SalI bands representing partial methylation being present. This plant was open pollinated in a greenhouse where no P-rr plants were present. Of five progeny plants from this ear, one gave a fully red ear, consistent with the possibility that this was a stable revertant to P-rr. Analysis of the second generation of progeny is under way.
In summary, the P gene in P-pr appears to be hypermethylated, and hypermethylation is linked to its patterned phenotype. In a discussion of the role of DNA methylation in epigenetic phenomena, Holliday defined epimutation as a mitotically heritable change in the methylation of a gene (Holliday, Science 238:163, 1987). Forward or reverse epimutations were defined as an increase or decrease in methylation, respectively. Based on this definition, the origin of P-pr may represent a forward epimutation. Furthermore, this example suggests that a change in the methylation of a gene can be heritable through meiosis, and can contribute to allelic diversity.
Figure 1. Genetic analysis of the patterned pericarp phenotype.
Figure
2. Hypermethylation in P-pr: A restriction map of the P-rr
gene is shown. A large direct repeat at the locus is represented by the
cross hatched box. Exons E1, 2 and 3 (horizontally striped boxes) make
up the functional transcript, and an alternate transcript arises from E1+2+4.
The probes used in the Southern analysis are shown above the map. Probe
15 detects the four repeats specific to the locus represented by the black
box, and probe 8B detects the large intron. SalI sites (S) are numbered;
E = EcoRI, B = BamHI. Leaf DNA from a P-rr/P-rr
plant is compared to leaf DNA from three sibling P-pr/P-ww
progeny after digestion with SalI. No differences are seen with
8B, controlling for partial digestion, but probe 15 detects a number of
higher MW bands. Three bands are detected for P-rr, of 1.2kb (two
comigrating bands), 3.0kb and 3.4kb. In P-pr, the higher MW bands
can be classed into partial hypermethylation (two bands of MW 4.2 and 4.6
from methylation of sites 4 and 8) and full methylation (two unresolved
bands of >10kb from methylation of sites 1, 2 and 4, and cutting at 5 and
a site to the left that is not shown, or methylation of 8 and 9, and cutting
at 7 and 10). Sites 3 and 6 are methylated in P-rr, and remain so
in P-pr. Sites 5 and 7 are unchanged in methylation. With seven
other enzymes, including HaeIII, HpaII, MspI, EcoRII
and PstI, probe 15 gave higher MW bands, while probe 8B showed no
differences These results indicate that the regions around the four filled
boxes are hypermethylated at both CpG and CpXpG in P-pr. However,
the intervening regions comprising much of the transcription unit appear
to be unchanged in methylation.
Return to the MNL 67 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}